3 Increase in Teretriosoma nigrescens with Prostephanus truncatus as the host
3.1 Breeding Prostephanus
truncatus and Teretriosoma
nigrescens
3.2 Population dynamics of Teretriosoma
nigrescens
3.3 The ability of Teretriosoma
nigrescens to reproduce in cultures of Prostephanus
truncatus on various substrates
3.4 The ability of Teretriosoma
nigrescens to reproduce on Prostephanus
truncatus populations after several months
on plant substrate
3.1 Breeding Prostephanus truncatus and Teretriosoma nigrescens
3.1.1 Breeding Prostephanus
truncatus
3.1.2 Breeding Teretriosoma
nigrescens
Preliminary experiments were set up to test the reproduction rate of T. nigrescens in varying predator/prey relationships. Taken into consideration here, were the age of the P. truncatus stock and, related to this, the varying profile of the stages of its development as prey organisms for the predator. The results of these preliminary experiments finally led to the following method for rearing T. nigrescens and P. truncatus.
3.1.1 Breeding Prostephanus truncatus
Mass breeding of the host, P. truncatus, was the basis for reproduction of the predator T. nigrescens. P. truncatus has been reared successfully at the Federal Biological Research Centre for Agriculture and Forestry (BBA), in Berlin at the Institute of Stored Product Protection since 1983. The populations for this purpose had been imported from Tanzania. Based on this method of breeding, approx. 500 P. truncatus imagines (this corresponds to a volume of about 5 ml) were placed on about 500 g shelled maize (trade name: "La Plata" from Argentina). The vessels used for breeding were jars with a volume of 2 litres and with wire gauze welded to their clip-on lids (Fig. 9A). After 4 - 5 weeks at 30 � 1�C and 75% � 5% r. h. without light, the F1 generation of the pest hatched. There were then about 2,000 to 3,000 adult specimens of P. truncatus in the jars which were used for further breeding, for experiments or to breed T. nigrescens.
3.1.2 Breeding Teretriosoma nigrescens
The initial population for breeding T. nigrescens was brought from Costa Rica in 1988. The breeding jars with P. truncatus set up as above were preincubated in climatic chambers without light at 30 � 1�C and 75% � 5% r. h.. Subsequently, 50 T. nigrescens imagines were inserted into each jar to lay their eggs. This corresponds to a predator/prey ratio of 1:10. A week later, the T. nigrescens adults were removed from the P. truncatus jars and were again available for further breeding.
Eight weeks (at 30 � 1�C and 75% � 5% r. h. without light) after the T. nigrescens imagines were put in with the host populations, approx. 70 150 F1 T. nigrescens imagines between the ages of 0 and 14 days could be removed. They were used for further breeding and for experiments.
A count of the adult F. specimens in 28 T. nigrescens breeding jars showed an average of 0.47 � 0.15 progeny/females and day. According to the results in paragraph 3.2, it was assumed that 50% of the T. nigrescens imagines in the P generation were female.
This method allowed mass breeding of T. nigrescens within a relatively short period. T. nigrescens and, linked to this, P. truncatus were bred so that freshly hatched specimens of the beetles were available at intervals of about 2 weeks.
3.2 Population dynamics of Teretriosoma nigrescens
3.2.1
Introduction
3.2.2
Material and method
3.2.3
Results
3.2.4
Conclusion
To be able to make any statements on the reproduction potential of a T. nigrescens population, a long-term experiment was carried out to determine the number of progeny a T. nigrescens female can produce during her life.
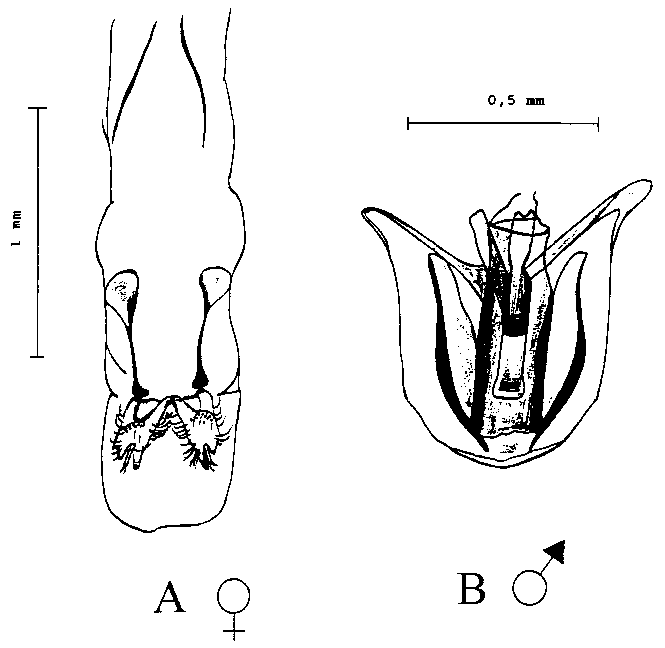
The basis for the investigation was the ability to define the sex of the T. nigrescens specimens. It is not possible to distinguish between male and female T. nigrescens imagines from their appearance. For this reason, the sex of the insects was defined by examining their genitalia after dissection.
Apart from this, the ratio of males to females within a T. nigrescens population was also defined.
To identify the sex organs, adult beetles were killed in 70% ethanol and attached to a slide with the underside upwards. This facilitated dissection of the genitalia of these extremely compact insects. Dissection was carried out at a magnification of 20 - 30 times. After removal, the genitalia were soaked in approx. 60% lactic acid (C3H6O3) for some minutes to make the surrounding tissue lighter. A comparison with illustrations from TANNER (1925) clearly showed which of the varying forms of genitalia came from a female.
To define the ratio of sexes within a T. nigrescens population, the process was repeated 10 times, each time with 100 T. nigrescens imagines from the breeding jars. The insects were all killed in 70% ethanol. Natural specimens were then pressed onto slides. The genitalia uncovered were then allocated to each sex under a binocular microscope.
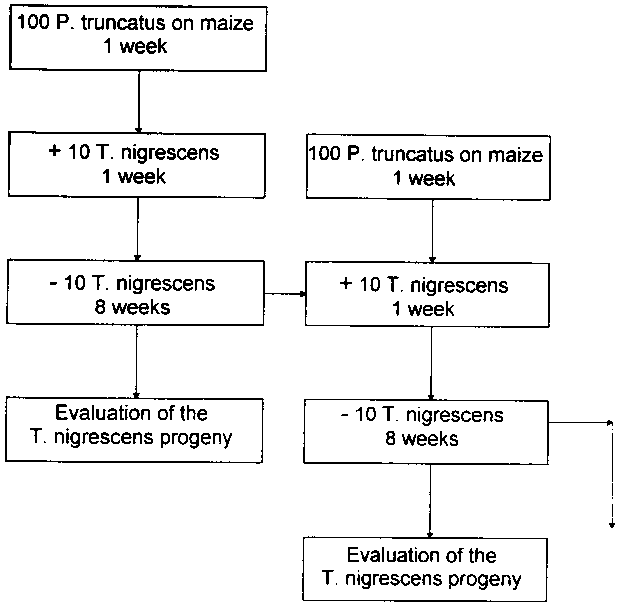
To find out the number of F. specimens a T. nigrescens can produce, P. truncatus populations on maize were selected as hosts. In 5 parallel experiments, lots of 100 P. truncatus imagines were put into 300 ml screw-top jars on approx. 150 g maize grains. These were then incubated in the climatic chamber for a week at 30 �1�C and 70% � 5% r. h. without light, so that when the predators were put in, eggs and young larvae of the hosts were already in the test jars. Finally, 10 T. nigrescens imagines of unknown sex and aged between 0 and 2 weeks were put into each jar.
The T. nigrescens females had the opportunity to lay their eggs on the P. truncatus broods for seven days in the experiments, which were continually subjected to the climatic conditions described during this time. Afterwards, the adult T. nigrescens specimens were removed from the host populations and, once more, put on 7-day-old P. truncatus broods.
To promote further development of the T. nigrescens eggs which had been laid, the host populations were incubated in the climatic chamber for another 8 weeks after the T. nigrescens imagines had been removed. The number of T. nigrescens progeny (imagines and larvae ready to pupate) in the P. truncatus brood was then counted (Fig. 2).
The initial ratio of predator to prey imagines was kept constant at 1:10 throughout the whole experimental period. If one of the 10 T. nigrescens imagines in the P generation died during the experiment, only 90 adult P. truncatus were used for the following host population. The quantity of maize was altered to approximately correspond to the extent of the P. truncatus population. However, less than 50 g maize was not used to make boring into the grains easier for the P. truncatus imagines.
{kind=link}
{kind=link}
The genitalia of the T. nigrescens imagines of the F1 generation which died during the experimental period were examined according to the method described to define their sex.
T. nigrescens imagines were put into new P. truncatus populations until examinations showed no progeny in the P. truncatus populations for at least 4 consecutive weeks.
At the end of the experiments, the sex of all the T. nigrescens imagines which were still alive was defined. After having found out how many females had been in each series of experiments, the number of progeny a T. nigrescens female had produced on average per day was calculated. For this, the mean values from four consecutive recordings in each series of experiments were worked out first. This corresponded to a period of investigation amounting to 4 weeks (28 days) in each case.
To summarise the results from all 5 series of experiments, the average was taken from these mean values. In this way, each data point in Fig. 4 derives from 20 single values.
The genitalia of a T. nigrescens male/female is illustrated in Fig. 3. The dissection of the genitalia from 1000 T. nigrescens imagines produced a sex ratio of almost 1:1 (52.0 � 2.5 males to 48.0 � 2.5 females).
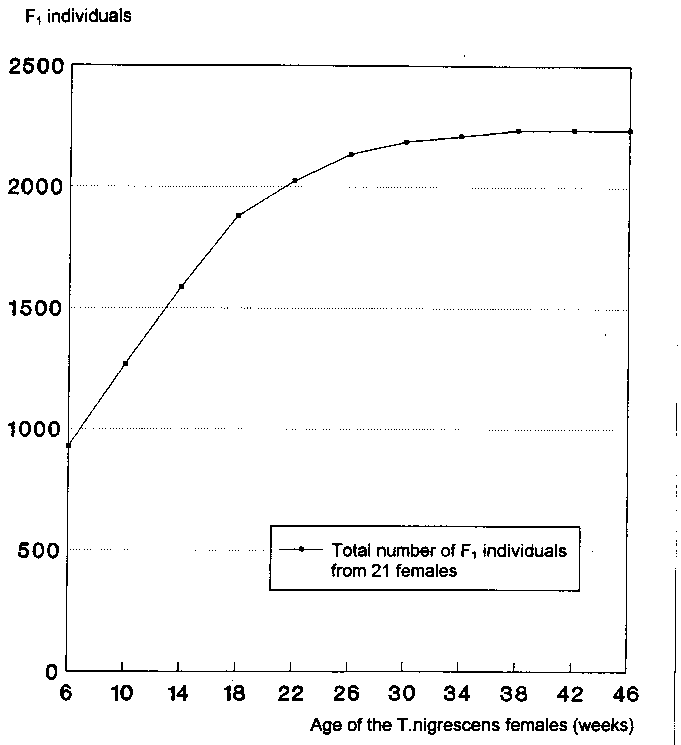
In 4 of the 5 series of long-term experiments were 4 female T. nigrescens in each case, in one series there were 5 females of the predator. Thus, in total, 21 of the 50 T. nigrescens imagines of the original P generation were females.
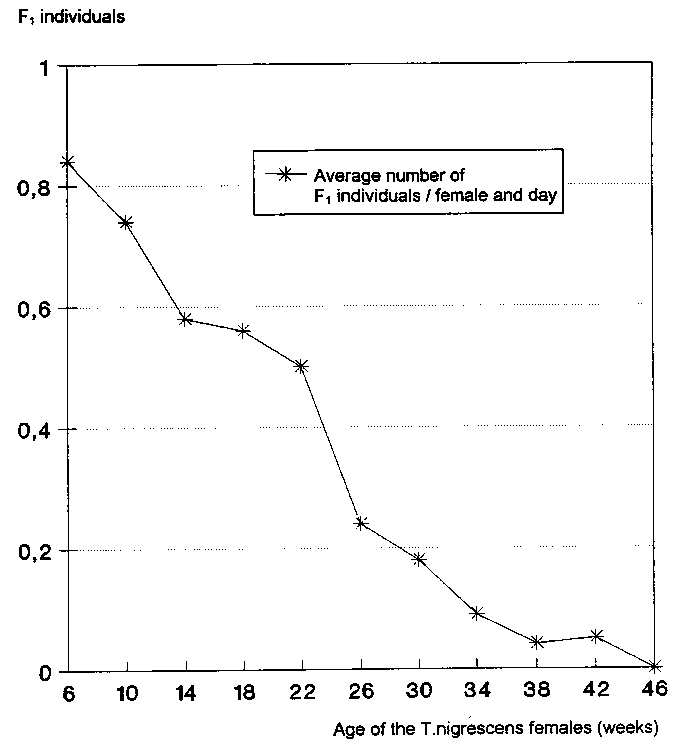
The number of progeny produced per day decreased as the females became older (Fig. 4). Female T. nigrescens of up to 6 weeks of age provided the maximum value of an average of 0.84 � 0.19 F1 specimens per female and day.
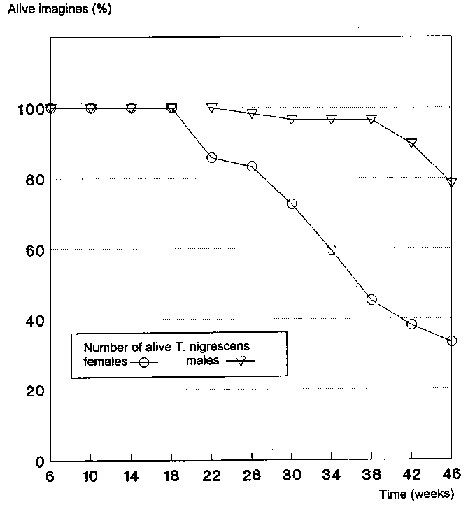
From an age of 18 weeks, the first T. nigrescens females began to die. The number of progeny in the P. truncatus populations fell up to this point by 33.3% to 0.56 � 0.08 F1 specimens per female and day. After 46 weeks, no more progeny could be determined for the 7 T. nigrescens females still alive (Fig. 5).
One T. nigrescens female was thus able to produce an average of 107 progeny during the course of her reproductive phase.
Under the conditions existing, the T. nigrescens females died earlier than the males (Fig. 6). Only one of the 29 males died within its first 38 weeks of life.
At the end of the experiment, 78.4% of the males and 33.3% of the females were still alive. Seen in total, 59.5% of the 50 T. nigrescens imagines initially inserted were still alive after 46 weeks (10.7 months).
Fig. 4: Number of T. nigrescens progeny in P. truncatus cultures depending on the age of the females
{kind=link}
{kind=link}
Fig. 6: Life expectancy of T. nigrescens imagines in P. truncatus cultures
{kind=link}
In this experiment, the number of F1 specimens was recorded which successfully developed from the T. nigrescens eggs into larvae able to pupate or into adults. Normally, not all eggs develop into adult insects. There are natural mortality rates within the development stages of individuals in a population which implies that there were more eggs laid by the T. nigrescens females than F1 specimens could be observed.
The following comparison of values shows that not only the "absolute" age of the female T. nigrescens can be a decisive factor in the number of offspring produced. The conditions under which the insects were kept before and whilst laying eggs have an effect on the "relative" age of the females: in the control experiments with T. nigrescens on plant substrate (see 5.2.1), the imagines were put together-with new P. truncatus at intervals of 4 weeks. Progeny could still be observed from the T. nigrescens females after 15 months.
When the T. nigrescens imagines were kept on maize without any hosts (see 3.4), 16.5-month-old T. nigrescens females could also produce progeny after being put together with P. truncatus populations. In contrast to this, 10.7 month-old T. nigrescens females were not able to produce progeny any more in these experiments.
The females presumably found optimum conditions for laying eggs in the constant new provision of P. truncatus cultures. Seen physiologically, the enormous amount of energy necessary for producing the eggs made the females age more quickly in comparison to the insects which had produced no progeny. This could possibly explain why the T. nigrescens females died relatively quickly in comparison to the males and also the high life expectancy of the insects on plant substrates.
It is improbable that the varying results on the ability to reproduce and on the lifetime of T. nigrescens are due to natural fluctuations among the individuals in a population. The results show that the age of the females used in experiments investigating the rate of egg-laying should be known and that the conditions for the insects should be taken into consideration.
3.3.1
Introduction
3.3.2
Material and methods
3.3.3
Results
3.3.4
Conclusion
So far, experiments on the ability of T. nigrescens to reproduce and its effect as a predator using P. truncatus as a host have been carried out on maize. Most storage pests however, do not rely, for example, on a particular substrate for breeding. P. truncatus is also able to reproduce on other substrates in addition to maize. For example, (B�YE, (1990) was able to show that P. truncatus reproduces on cassava, sorghum and rice.
The following experiments were to examine whether T. nigrescens could suppress growth of a P. truncatus population on substrates other than maize and reproduce on these.
The substrates used were cassava, wheat, sorghum, peas and bran (wheat bran mixed with glycerine, glucose and yeast has been ideal for breeding many species of Coleoptera and Lepidoptera storage pests).
Per series of experiments, 10 x 300 ml, glass vessels were filled half-full with 150 g wheat, sorghum or peas, or 100 g cassava, or 50 g wheat bran. Subsequently, 100 P. truncatus imagines were put into each vessel. After an incubation period of 7 days in the climatic chamber at 27 � 1�C and 75% � 5% r. h. and without light, 10 young T. nigrescens imagines were added to half the vessels in each series (n=5). The remaining experiments without predators served as controls. Evaluation was carried out eight weeks after inserting the adult T. nigrescens. The number of P. truncatus imagines and the number of T. nigrescens progeny in the experiments were counted. The comparative values used were from two series of experiments with P. truncatus on maize. Since P. truncatus is also described as being a pest on cassava, two sets of experiments were also carried out with dried cassava roots from Togo. The statistical evaluation was carried out using the t-test according to RENNER (1981). To do this, the arithmetic mean (MOO) with the standard deviation (SD) was calculated from the number of host imagines in each of the 5 experiments without predators and with T. nigrescens. A subsequent comparison of these mean values (t-test) clearly showed whether evidence of an influence of T. nigrescens on the growth of the beetle populations existed or not.
If the mean values varied with an error probability
| of > 5% | the influence of T. nigrescens was considered to be non-existent (N) |
| of � 5% | the influence of T. nigrescens was considered to be probable (PR) |
| of � 1% | the influence of T. nigrescens was considered to be significant (SI) |
| of � 0.1% | the influence of T. nigrescens was considered to be highly significant (HS). |
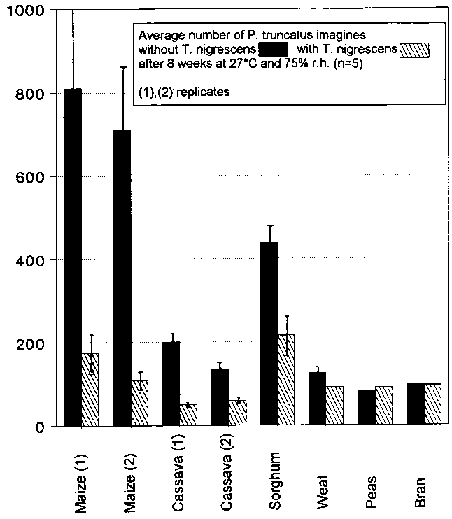
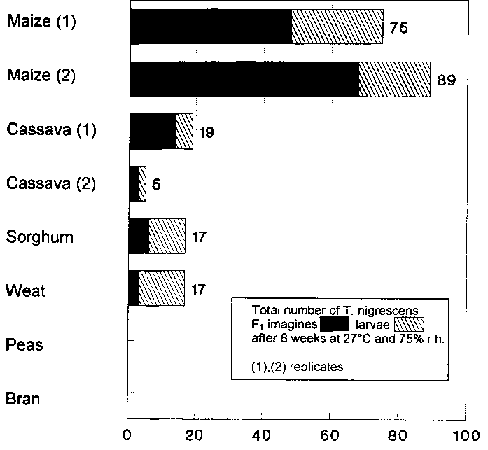
P. truncatus was able to reproduce most successfully in the experiments on maize (Tab. 1). The host populations grew to seven and eight times the size of the initial populations of 100 imagines. In the experiments with T. nigrescens at 78% in the first series and 85% in the second, there were highly significantly less P. truncatus imagines than in the controls without the predator. In the first experiment, 48 imagines and 27 larvae, in the repetition a total of 68 F. imagines and 21 larvae of T. nigrescens could be found.
P. truncatus was only able to reproduce weakly on cassava. In one of the two series, an average of around 200 imagines was observed, in the other, 110 -150 per experiment (Tab. 1). T. nigrescens was able to achieve a highly significant reduction of 75% and 55% respectively in the number of adult hosts (Fig. 7). In these P. truncatus cultures, T. nigrescens was hardly able to reproduce. Only 19 progeny of the predator could be found.
Table 1: Influence of 10 T. nigrescens on the development of P. truncatus cultures on various substrates and the ability of the predator to reproduce on these cultures
100 P. truncatus imagines
on various substrates; evaluation after 8 weeks at 27�C and 75%
r.h. (mw�sd, n=5)
*: all P. truncatus were dead; (1), (2): replicates
SI, HS: varying from controls without T. nigrescens
significant (p � 1%), highly significant (p� 0,1%)
| Experiment | Adult P. truncatus |
Progeny T. Nigrescens |
Substrate |
| P.trun. (1) | 810.2 � 190.6 | Maize | |
| P.trun. + T.nigr. | 175.6 � 52.3 HS | 15.0 � 4.9 | |
| P. trun (2) | 710 8 � 153 2 | Maize | |
| P. trun + T.nigr. | 109 6 � 25 1 HS | 17.8 � 2.2 | |
| P.trun.(1) | 200.4 � 19.0 | Cassava | |
| P.trun. + T.nigr. | 50.4 � 7.8 HS | 3.8 � 2.0 | |
| P.trun.(2) | 132.6 � 17.4 | Cassava | |
| P.trun. + T.nigr. | 59.4 � 12.0 HS | 1.0 � 0.7 | |
| P.trun. | 438.0 � 40.0 | Sorghum | |
| P.trun. + T.nigr. | 215.8 � 50.6 HS | 3.4 � 2.2 | |
| P.trun. | 125.2 � 16.0 | Wheat | |
| P.trun. + T.nigr. | 91.2 � 8.4 SI | 3.4 � 2.8 | |
| P.trun. | 81.6 � 2.6* | Peas | |
| P.trun. + T.nigr. | 90.6 � 4.8* SI | 0 | |
| P.trun. | 97.6 � 1.8 | Bran | |
| P.trun. + T.nigr. | 96.2 � 1.3 | 0 |
{kind=link}
{kind=link}
P. truncatus reproduced relatively well on sorghum (Tab. 1). There were 380 - 450 P. truncatus imagines in each experiment. The number of adult hosts was reduced highly significantly by 51% by T. nigrescens in comparison to the controls (Fig. 7). In these cultures, a total of 6 F1 imagines and 11 larvae of T. nigrescens were found.
Wheat was a very unsuitable substrate for P. truncatus to breed upon. In experiments without the predator, there were only 110 - 150 imagines per vessel (Tab. 1). With T. nigrescens, there was a significant number of 27% fewer adult P. truncatus (Fig. 7). In total, 3 F1 imagines and 14 larvae of the predator could be found.
There was no evidence of reproduction of P. truncatus, nor could any progeny of P. nigrescens be found in the tests on peas or bran. No reductive influence of T. nigrescens could be determined on the number of adult hosts (Fig. 7). In the experiments with T. nigrescens on peas, significantly more P. truncatus imagines could even be found than in the controls.
On all the substrates on which P. truncatus was able to reproduce (maize, cassava, sorghum and wheat), offspring of T. nigrescens could also be observed.
Reproduction of the predator (number of parental insects inserted/number of F1 specimens > 1) only took place within the experimental period of 8 weeks, when the host populations were kept on maize. Presumably, more predators were able to develop in these P. truncatus populations than in the other host populations due to the large supply of food.
T. nigrescens was also able to reproduce in the P. truncatus populations when the medium used as a substrate for breeding was less suitable for P. truncatus and thus only a few prey organisms were available, e.g. on cassava and on wheat. T. nigrescens suppressed growth of the P. truncatus populations on all substrates used for breeding the pest. Thus, successful control of P. truncatus using T. nigrescens is not only possible on maize but also on other substrates infested by the pest.
3.4.1
Introduction
3.4.2
Material and method
3.4.3
Results
3.4.4
Conclusion
Evidence of reproduction of T. nigrescens on plant substrate without hosts could not be found (5.4). The following experiments were carried out to test whether this was a pause in reproduction or an irreversible loss of the ability to reproduce.
After 9.5 months, 10 beetles (Tab. 8) were taken from each of some of the experiments described under 5.2 (n=11) where a lot of T. nigrescens were still alive (maize, cassava, beans, sorghum, wheat, oats, peanuts, almonds and rice), and put into 7-day-old P. truncatus cultures (100 adults on approx. 150 g maize in 300 ml glass vessels). As a control, young T. nigrescens bred in vessels were collected and also put into new host populations (n=5). Another control consisted of P. truncatus populations without any influence from the predator (n=5). After an incubation period of 8 weeks in a climatic chamber at 27 � 1�C and 75% � 5% r. h. without light, the number of T. nigrescens progeny and the number of adult P. truncatus were counted.
After 16 months, a second experiment was carried out. The beetles were taken from tests with maize, cassava, sorghum, wheat and oats (n=6). Evaluation took place after 12 weeks.
After a reproduction pause of 9.5 months, T. nigrescens was able to again reproduce on the populations of its host, P. truncatus After approx. 2 months, there was an average of 9.5 � 5.0 progeny (imagines and larvae) of T. nigrescens. In connection with this, the beetles retained their ability to suppress the growth of the host populations. The controls without T. nigrescens contained an average of 810.2 � 190.6 adult P. truncatus whilst in the jars with the predators, only 250.7 � 90.4 hosts could be counted, corresponding to a reduction of 69%. The average number of T. nigrescens progeny in the tests with the young control specimens from the breeding jars was a little higher at 15.0 � 4.9. In these experiments, an average of 175.6 � 52.3 adult P. truncatus were counted. In comparison to the controls without any T. nigrescens specimens, this corresponds to a reduction of 78%.
Even after 16 months on plant nutrient media, T. nigrescens was able to reproduce on P. truncatus cultures and suppress the growth of the pest populations. After 3 months there was an average of 30.7 � 12.2 progeny of T. nigrescens and 158.8 � 74.5 adult P. truncatus in the experiments. The unaffected P. truncatus populations grew to an average of 653.3 � 193.6 imagines. This corresponds to a reduction by T. nigrescens of around 76%.
Cannibalism must be seen as an explanation for the results determined. Due to insufficient nourishment, the T. nigrescens imagines eat their own eggs. The eggs are not safe until there are sufficient other prey organisms available. (See also 5.2 concerning this.)
Another theory is that the purely vegetarian nourishment of T. nigrescens induces an irreversible pause in reproduction. The lack of animal protein, cholesterol or other essential nutrients, or with these only being present in plant nutrient media in insufficient quantity, could lead to inhibition of oogenesis. As soon as the nutrient is available from animal nourishment, the maturing process of the eggs begins again.
It can be assumed that the T. nigrescens females, when they were placed in the jars with the plant substrates, had already mated. The sperm of the males enters the receptaculum seminis, a kind of store for the sperm, through the sex organ of the female. The eggs which are ready to be laid are fertilised in the ovary tubes when passing the receptaculum (WEBER & WEIDNER 1974). Often, only one mating is sufficient to fertilise all the eggs maturing in a female. Inhibition of spermatogenesis on plant substrate thus often plays a more subordinate role.
The predators are quickly able to lay fertilised eggs again. The T. nigrescens imagines kept on coarse maize meal for 3 months were ready and able to lay eggs again in the P. truncatus cultures within 7 days, from which adult T. nigrescens developed.