 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Use of Trees by Livestock (NRI, 1994, 160 p.)
Use of Trees by Livestock: Prosopis
N.J.L. Clinch, l.J. Bennison and R.T. Paterson
Acknowledgements
© Crown copyright 1993
The Natural Resources Institute (NRI) is an internationally recognized centre of expertise on the natural resources sector in developing countries. It forms an integral part of the British Government's overseas aid programme. Its principal aim is to alleviate poverty and hardship in developing countries by increasing the productivity of their renewable natural resources. NRI's main fields of expertise are resource assessment and farming systems, integrated pest management, food science and crop utilization.
NRI carries out research and surveys; develops pilot-scale plant, machinery and processes; identifies, prepares, manages and executes projects; provides advice and training; and publishes scientific and development material.
Short extracts of material from this publication may be reproduced in any non-advertising, nonprofit-making context provided that the source is acknowledged as follows:
Clinch, N.J.L., Bennison, J.J., and Paterson, R.T.
(1993) Use
of Trees by Livestock 1: Prosopis.
Chatham, UK: Natural Resources Institute.
Permission for commercial reproduction should be sought from:
The Head, Publications and Publicity Section, Natural Resources Institute, Central Avenue, Chatham Maritime, Kent, ME4 4TB, United Kingdom.
No charge is made for single copies of this publication sent to the governmental and educational establishments, research institutions and non-profitmaking organizations working in countries eligible for British Government Aid. Free copies cannot normally be addressed to individuals by name but only under their official titles.
Natural Resources Institute ISBN: 0 85954 332 3 Price £2.00 LG1
This booklet is printed on paper produced using the residues from sugar-cane processing.
Foreword
The importance of trees and shrubs in feeding animals in the tropics and sub-tropics has long been recognized by livestock owners. In arid areas where the growth of herbaceous plants is limited by lack of moisture, leaves and edible twigs of trees and shrubs can constitute well over 50% of the biomass production of rangeland. At high altitudes, tree foliage may provide over 50% of the feed available to ruminants in the dry season, branches being harvested and carried to the animals. Even in regions of higher rainfall where grass supplies the major proportion of the dry matter eaten by ruminants, tree leaves and fruits can form an important constituent of the diet, particularly for small ruminants.
In the last two decades interest in planting trees as a source of feed for livestock has been encouraged by workers in research and development, but in contrast to the hundreds of indigenous species which are used as fodder, attention has focussed on a limited number of introduced species. Thus there are many publications reporting the chemical composition of Leucnaena leucocephala leaves and suggesting management strategies for utilization of the tree for fodder, but it is more difficult to find information on alternative genera which might be equally, or more appropriate.
The aim of this series of publications is to bring together published information on selected genera of trees which have the potential to increase the supply of fodder for ruminants. Each booklet summarizes published information on the fodder characteristics and nutritive value of one genus, with recommendations on management strategies, where available. Further, since the leaves of woody species frequently contain secondary compounds which may have an anti-nutritional, or toxic, effect, a separate booklet summarizes the effects of a number of these compounds. It is hoped that the booklets will provide useful resource material for students, research and extension workers, interested in promoting the use of trees as a source of fodder for ruminants.
Margaret Gill Livestock Production Programme
Genus Prosopis
Family LEGUMINOSAE
Subfamily MIMOSOIDEAE
Tribe
ADENANTHEREAE
Subtribe FABACEAE
|
Principal species |
Prosopis africana |
| |
Prosopis alba |
| |
Prosopis articulata |
| |
Prosopis caldenia |
| |
Prosopis chilensis |
| |
Prosopis cineraria |
| |
Prosopis flexnosa |
| |
Prosopis glandulosa |
| |
Prosopis juliflora |
| |
Prosopis nigra |
| |
Prosopis pallida |
| |
Prosopis tamarugo |
| |
Prosopis velutina |
|
Main common names |
Mesquite (North America) |
| |
Algarrobo (South America) |
| |
Khejri (India) |
Summary
Prosopis includes species that are extensively used, or have great potential in arid and semi-arid regions for a multitude of purposes. There are, however, constraints to their inclusion in agroforestry systems. While the fruits are valuable feeds for man and his livestock, anti-nutritive factors, particularly the presence of tannins and alkaloids, have been reported and some species have the potential to become noxious weeds. Nevertheless, remarkable tolerance of heat, drought and soil salinity, the ability to recover from severe defoliation and a wide range of uses are attributes that place the genus amongst the most promising plants for reafforestation and development of harsh, tropical regions.
Description and distribution
Prosopis is a genus of sub-tropical trees and shrubs, many of which are armed with prickles, or straight, stout spines. The leaves are bipinnate, tending to drop under the influence of severe climatic stress. The flowers are small and greenish white in colour. Pods, which show little tendency to shatter, are often formed in clusters and contain a sweet pulp surrounding each seed. This is attractive to both humans and livestock. The wood is close-grained and heavy and the root system extensive. The genus is well adapted to arid and semi-arid regions, where it is favourably regarded for rehabilitation of land that would otherwise remain economically worthless (NAS, 1979).
Species delineation is confused by synonymy and variable forms, but 44 species are generally recognized. While centres of origin appear to be in South America and lndia, the genus is now widespread throughout sub-tropical North and South America, Asia and Africa but uncommon in Europe and Australia (Allen and Allen, 1981).
Within North America, where Prosopis spp. are commonly known as mesquite, P. glandulosa and P. velutina have potential to become devastating weeds. These species are sometimes confused with P. juliflora, which shows much promise as a multipurpose tree for arid regions. In some areas, mesquite leaves and pods are major sources of forage for a range of livestock and indigenous animals during the dry season (Skerman et al., 1988).
In South America, P. chilensis is widespread, extending southwards into Chile and Argentina. With other species such as P. alba, P. flexuosa and P. nigra, it is known by the general term algarrobo, and is considered to be a vital part of agrosilviculture and pastoral land use systems in the semi-arid regions of Argentina (Marmillon, 1986).
P. cineraria (khejri) is native to Arabia, Iran, Afghanistan, Pakistan and India and is worshipped by the people of the Thar desert, who value it as a famine food (Harris et al., 1989). It has been described variously as the most important top-feed (browse carried at a height well above ground level) for all livestock in the desert areas of Asia (Bohra, 1980), and as an unexploited treasure of the Thar desert (Jatasara and Paroda, 1981).
P. africana is native to parts of North, West, Central and East Africa but is mainly used as a durable hardwood for furniture and construction and for charcoal (Dalziel, 1948; Booth and Wickens, 1988). P. ch�lensis has been introduced into West Africa and P. juliflora is now naturalized in India, West Africa and the Sudan, where it prefers soils with a shallow water table, or a coastal climate (Le Houerou, 1980).
Fodder Characteristics
Prosopis is recognized as an important source of fodder for all livestock species in arid and semi-arid environments, although there are conflicting reports regarding the palatability and nutritive value of the leaf material. Representative data for proximate and fibre analyses are shown in Table 1. In India, Parthasarathy (1986) found that leaves of P. cineraria were well accepted by weaner kids (daily voluntary dry matter (DM) intake of up to 4.3% of bodyweight), but that animal performance was limited by low digestibility, possibly due to excessive lignification of the twigs and leaves. Lyon et al., (1988) considered that the leaves of P. alba, P. articulata, P. chilensis, P. nigra, P. velutina and P. velutina var. Ruby were suitable sources of forage. Conversely, NAS (1979) suggested that the major contribution of the genus was in terms of pod production and that where leaves were consumed, this could be seen as an occasional bonus. This view appears to be shared by both Ibrahim and Gaili (1985), and Abdelgaabar (1986), who classed P. chilensis foliage as unpalatable. The latter author went so far as to suggest that supplementation with this forage source should consist entirely of pods. Leaf acceptability varies with species and possibly also with geographic location. Le Houerou (1980) described P. juliflora as moderately palatable, while Toutain (1980) noted that P. chilensis was not appreciated greatly by livestock and P. africana was occasionally appealing. Nevertheless, Le Houerou, (1980) recommended lopping systems of management for Prosopis spp. and classed P. cineraria as an important source of top-feed during lean months in the more arid parts of Asia. In the Rajasthan area of India, leaves of P. cineraria are collected in the period from February to March for feeding later in the year (Bhandari et al., 1979). The leaves have a relatively high crude protein content (15-16%), although the digestibility is usually low.
Prosopis pods have long been recognized as feed for both human and livestocks. Harden and Zolfaghari (1988) report that whole pods contain 91% crude protein, 13-31% sucrose and 17-31% crude fibre (see Table 1).
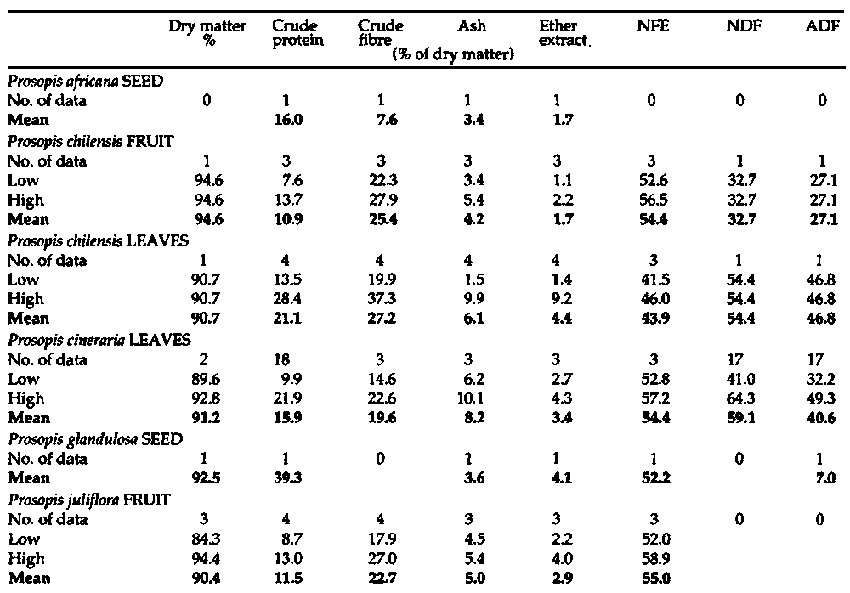
Table 1

Table 2
Pods are produced hanging on small stalks in clusters of up to 12. Usually about 20 cm long, they are either flat or coiled into a spiral. They contain several seeds embedded in a sweet, dry, yellow pulp. The amount of pulp, which is principally sucrose, varies between individual trees. NAS (1979) noted that cattle, sheep, mules, donkeys, goats and wildlife all consume them avidly. Pods may be produced twice per year, but estimates of the feeding efficiency and digestibility vary widely.
While steers on over-grazed pastures in semi-arid areas of South America lost liveweight, supplementing their diet with mature pods of P. caldenia with a DM digestibility of 63.8% resulted in weight gains (Menvielle and Hernandez, 1985). Pods of P. juliflora were successfully used to replace wheat bran at varying levels in diets for crossbred calves and lambs. Incorporation affected neither nutrient digestibility of rations nor balances of nitrogen, calcium and phosphorus. No significant differences were reported either for average daily gain, feed intake, feed efficiency or carcass characteristics (Rao and Reddy, 1983). In contrast, when desert Sudanese goats were fed on crushed pods of P. chilensis, cottonseed meal and wheat bran in differing proportions, intake, liveweight gain and dressing-out percentage all decreased linearly with increasing proportion of pods. A bitter taste was noted, which may have contributed to the low intake (Ibrahim and Gaili, 1985).
It is generally accepted that P. tamarugo fruits are a reasonably good feed for ruminants, but in two trials with eight Merino sheep in Chile, individual estimates of digestibility ranged from 37.7 to 60.1% for crude protein and from 32.8 to 46.3% for crude fibre (Habit et al., 1981). The wide variation between animals could be due to some seed passing undigested through the gut. Punj (1988) saw this as a major problem and recommended development of suitable processing technologies.
Published analyses of the mineral and amino acid composition of Prosopis tend to be restricted to those species which are eaten by humans in the Americas. Some data are presented in Table 2. They appear to show a generally satisfactory profile, although total sulphur containing amino acids could be limiting (Felker and Bandurski, 1979; Harden and Zolfaghari, 1988).

Table 2 - Amino acid composition of
Prosopis seeds and
pods
Anti-nutritive factors
Harden and Zolfaghari (1988) reported that green and ripe Prosopis pods were used as food for both humans and animals. Gohl (1981), however, stated that green pods were bitter, with little feed value and that only ripe pods were relished by stock. Gohl went on to state that stock poisoning had occurred when P. juliflora pods were eaten after being exposed to rain. NAS (1979) noted that cattle fed on an exclusive diet of pods of P. pallida and P. glandulosa became malnourished and developed jaw and tongue troubles which caused difficulty in chewing and resulted in weight losses. The anti-nutritive factors in pods are not well understood but it would seem prudent to balance a pod-based diet with other feedstuffs where possible.
There is some evidence that certain parts of P. africana contain the alkaloids prosopine and prosopinine (Booth and Wickens, 1988), although it is not certain that these compounds have an antinutritive effect on ruminants. Dalziel (1948) reported that the bark of P. africana trees in east Sudan contained 14-16% tannin, capable of giving a reddish brown colour to both leather and cloth. Tannin is recognized as an anti-nutritive factor when fed to ruminants at concentrations above about 5% (McLeod, 1974).
Tannins are also present in the leaves of certain Prosopis spp. including P. cineraria, P. juliflora and P. glandulosa and there are a number of reports of the negative effects of tannins on DM and crude protein digestibility (Bhandari et al., 1979; Kumar, 1983; Upadhyaya, 1985; Lyon et al., 1988 and Holechek et al., 1990). Kumar (1983) observed that low crude protein digestibility of P. cineraria had been attributed to the presence of high levels of tannins. The low in vitro digestibility of crude protein (32.8% for a crude protein content of 13.7%) appeared to be due to the presence of condensed tannins, estimated with a vanillin-HCl assay to be 160 mg/g, with a protein precipitating capacity of 10.6%. Further reports of low digestibility values for crude protein (average 30.2%) were attributed to the high content of tannins and structural carbohydrates in the leaves. Lyon et al., (1988) negatively correlated in vitro digestibility with phenolic content of various species of Prosopis. Livestock deaths have also been linked with consumption of Prosopis leaves with a high tannin content (Kumar, 1983).
Thus, especially with P. cineraria, it is clear that Prosopis, one of the most abundant and important top-feeds of desert regions, is both poorly digestible and occasionally causes stock losses. There are a number of possible explanations for these observations. Firstly, the levels of tannins in the leaves of Prosopis vary considerably. Sehgal (1984) reported tannin levels of 2.18%, measured by the Folin-Denis test, while in an examination of 15 individual trees, Joshi et al., (1985) found levels ranging from 10.6 to 25.3%. The latter authors did not record the analytical procedures used to determine the tannin contents and it may be that their data are not directly comparable with those of Sehgal (1984). Nevertheless, they serve to demonstrate extreme variability between the foliage from individual trees. Newly emerging leaves had a higher tannin content than older ones, showing greater phytochemical protection for the structures which are of most importance in plant survival. Secondly, environment and stress on the plant are thought to play a part in tannin concentration, although Vaithiyanathan and Singh (1989), while noting seasonal changes in the tannin contents of top-feeds in the arid region, failed to relate this to temperature variation. Lastly, Prosopis is not necessarily suitable as a sole feed for livestock. Some of the in vivo digestibility trials reported in the literature have used Prosopis as the sole feed for small ruminants, but in most on-farm situations, it is likely that animals would self select, or that the fodder would be fed in combination with other feeds. This would dilute the toxic principle and mitigate the more extreme effects.
Management
While Prosopis spp. have been traditionally valued in their centres of origin and have been introduced into many new areas, particularly in the arid and semiarid regions of the world, little is known about appropriate browse management practices to optimize animal productivity. The genus has a recognized capacity for recovery from severe injury such as lopping (Booth and Wickens, 1988), but lopping is not necessarily the best way to achieve optimum sustainable productivity of high quality forage, especially in those species where the foliage is less palatable than the fruit.
In the absence of comparative data, the following establishment and management practices employed with P. tamarugo in a reafforestation programme in the Tamarugal Pampa sector of the Atacama desert in Chile (Habit et al., 1981), are presented as practical examples of techniques which may be suitable for other species of Prosopis.
Seed production
There are wide differences between individual trees in terms of phenotypic characteristics. Selection of the seed producers is, therefore, of great importance. The mature pods are ground in a stone mill set at 4 mm and clean seed is obtained by sieving and flotation. Insects attack seed if it is not treated with 0.2% Aldrin solution or an equivalent, prior to storage.
Seed germination
Scarification is necessary to promote germination. Sulphuric acid has been used in Chile, although other authors suggest scalding, or treatment with giberellic acid (Booth and Wickens, 1988).
Seedling production
A potting mixture of two parts of soil to one part of sheep manure has been used to raise seedlings. Fumigation with methyl bromide, or treatment with fungicides prior to sowing reduces seedling mortality. Seeds are sown to a depth of 1.5 cm and carefully watered to avoid accumulation of water in the lower level of the soil. In pots or plastic bags 30 cm deep, seedlings are grown for a period of three to five months, when they are 8 to 10 cm tall. Roots grow faster than shoots and care must be taken to avoid them growing out of the bags.
Planting out
While spacings ranging from 6 x 6 m to 13 x 13 m have been tested, current plantings are carried out either at 10 x 10 m in square, or at the much lower density of 15 x 15 m in triangular formation. Closer spacing results in faster cover but this is offset by higher costs, particularly in watering, which is of vital importance in the desert environment. Within this range of tree densities, closer spacing produces faster canopy closure but appears to have little effect on forage yield of the mature forest.
Pruning
The abundant growth of thorny, basal branches necessitates pruning at between four and six years of age, to allow animal access to the forage which falls from the trees. In Chile, this is done by hand, using a sickle.
Utilization
Animals are allowed free access to the established area once the first crop of pods has fallen, about seven years after planting. All classes of livestock tend to pick up fallen leaves and pods, rather than to browse the growing herbage.
Productivity
Productivity of P. tamarugo is linked to age, plant spacing, depth and quality of groundwater and land preparation prior to planting. In Chile, trees reach their mature heights of about 10 m at some 24 years of age. Fruit production starts at about seven or eight years and increases steadily with tree height. After year 24, it continues to increase at a declining rate until it reaches its maximum at about year 35. Leaf production starts earlier but also increases to about year 35, by which time annual production of forage DM is about 190 kg/tree (Robertson, 1980), split equally between fallen leaves and fruits. Estimates of available forage related to the area of foliar projection on the soil surface range from 0.75 to 3.4 kg DM/m2 for young (10 years, Robertson, 1980) and mature (30 years, Habit et al., 1981) trees, respectively. Conservative estimates would suggest that in a relatively closed forest, yields of 10 t/ha DM would not be an unrealistic goal (Felker and Bandurski, 1979), and that this quantity would support about 12 sheep/ha on a year-round basis. Felker et al., (1984) indicated, however, that there was considerable variability in pod production, both between and within accessions, suggesting scope for selection and possibly even breeding to increase forage productivity.
Allernative uses
The benefits to be derived from the nitrogen-fixing ability of leguminous plants are well known and species of Prosopis are no exception. They are usually found growing in soils with poor nitrogen status where their leguminous nature allows them to thrive. Members of the genus have a variety of uses and, indeed, P. cineraria has been classed as a genuinely multipurpose tree (Leakey and Last, 1980) providing firewood, timber, fodder, soil improvement, shelterbelts, edible fruit, medicines, bee forage, sand dune stabilization and extractable tannins and edible gums.
The pods are an important source of human food. In the semi-desert regions of S. America, mature Prosopis pods are used to prepare a sweet, floury paste considered a valuable food, particularly for children. The sugary pulp from the pods can be fermented and distilled to produce ethyl alcohol. Green and ripe pods of P. cinerar�a are consumed in India. The green pods are used in curry dishes or are dried and preserved, while the pulp of mature pods is eaten by children (Harden and Zolfaghari, 1988).
The wood is highly valued, being compact, closegrained, heavy, and resistant to borers, termites and general decay. It is used for implement handles, walking sticks, floors, posts, barrels, cabinets and larger constructions. All species are used as firewood and Prosopis is renowned as an excellent, slow-burning charcoal (Allen and Allen, 1981; Habit et al., 1981; Booth and Wickens, 1988).
The bark of various species contains substantial quantities of mesquite gum, which resembles gum arable, has emulsifying properties and is an excellent mucilage. Tannins are also present in concentrations that justify commercial extraction (Allen and Allen, 1981).
Crane (1975) included Prosopis spp. amongst the very drought-tolerant, tropical plants in her classification of important world honey sources. The nectar gathered from these trees is purported to yield a honey of superior flavour (NAS, 1979), while the bark can be used to make beehives (Booth and Wickens, 1988).
In Africa, the wood of P. africana was traditionally credited with soporific properties (Dalziel, 1948), while the macerated leaves were thought to ensure male fertility (Uphof, 1968). Nearly all parts of the tree are used in local medicines (Booth and Wickens, 1988). Young roots are used as a diuretic and to treat dysentery, while the vapour from boiling them is used against bronchitis and to control vermin. The bark is used to make a mouthwash to alleviate toothache and an eyewash to treat opthalmia. Dried, powdered bark is used against leprosy, orchitis, rheumatism, dermatosis and fever and to heal wounds, while the leaves are used to treat head, eye, ear and tooth troubles, or as a bath for the treatment of migraine and vertigo. Broun and Massey (1929) reported that dried and crushed pods or fruit husks of P. africana were used as a fish poison.
References and further reading
ABDELGAABAR, A.I. (1986) 1. Proximate composition of mesquite, Prosopis chilensis (Molina) Stuntz, pods, seeds and leaves. 2. Digestibility trials. Pamphlet No.2, Prosopis Project. Sudan: Forestry Research Centre.10p.
ALLEN, O.N. and ALLEN, E.K. (1981) The Leguminosue: A Source Book of Characteristics, Uses and Modulation. Madison, USA: University of Wisconsin Press. 812p.
BHANDARI, D.S., GOVIL, H.N. and HUSSAIN, A. (1979) Chemical composition and nutritive value of khejri (Prosopis cineraria) tree leaves. Annals of Arid Zone, 18(3):170-173.
BOHRA, (1980) Nutrient utilization of Prosopis cineraria (khejri) leaves by desert sheep and goats. Annals of Arid Zone, 19(1/2):73-81.
BOOTH, F.E.M. and WICKENS, G.E. (1988) Non timber uses of selected arid zone trees and shrubs in Africa. FAO Conservation Guide 19. Rome: FAO.
BROWN, F.A. and MASSEY, R.F. (1929) Flora of the Sudan. London: Sudan Government Office.
CRANE, E. (1975) The plants honey comes from. Chapter 1 pp 3-76. In: Honey, a Comprehensive Survey. Crane, E.(ed.) London: Heinemann. 608p.
DALZIEL, J.M. (1948) The Useful Plants of West Tropical Africa. London: Crown Agents for the Colonies. 612p.
DUKE, J.A. (1981) Handbook of Legumes of World Economic Importance. New York and London: Plenum Press.
FELKER, P. (1979) Mesquite - An all purpose leguminous arid land tree. In: New Agricultural Crops. RITCHIE, G.A. (ed.) Proceedings, Symposium of American Association of Agricultural Science. Boulder, USA: Westview Press. 38:89-132.
FELKER, P. and BANDURSKI, R.S. (1979) Uses and potential uses of leguminous trees for minimal energy input agriculture. Economic Botany, 33(2):172184.
FELKER, P., CLARK, P.R., OSBORN, J.F. and CANNELL, G.H. (1984) Prosopis pod production Comparison of North American, South American, Hawaiian and African germplasm in young plantations. Economic Botany, 38(1):36-51.
GOHL, B. (1981) Tropical Feeds: Feed Information Summaries and Nutritive Values. FAO Animal Production and Health Series No.12. Rome: FAO. 529p.
GUPTA, T. (1983) The economics of tree crops on marginal agricultural lands with special reference to the hot arid region in Rajasthan, India. International Tree Crops Journal, 2(2):155-194.
HABIT, M.A., CONTRERAS, D.T. and GONZALEZ, R.H. (1981) Prosopis tamarugo: Fodder Tree for the Arid Zones. FAO Plant Production and Protection Paper No 25. Rome: FAO.
HARDEN, M.L. and ZOLFAGHARI, R. (1988) Nutritive composition of green and ripe pods of honey mesquite (Prosopis glandulosa, Fabaceae) Economic Botany, 42(4):522-532.
HARRIS, P.J.C., GATES, P., SOARES, H. and
THEISEN, P. (unpublished) Tree and Shrub Legumes for Arid and Semi-arid areas. Ryton-on Dunsmore, Coventry, UK: Henry Doubleday Research Organisation.
HOLECHEK, J.L., MUNSHIKPU, A.V., SAIWANA, L., NUNEZ-HERNANDEZ, G., WALLACE, J.D. and CARDENAS, M. (1990) Influences of six shrub diets varying in phenol content on intake and nitrogen retention by goats. Tropical Grasslands, 24:93-98.
IBRAHIM, A. and GAILI, E.S. (1985) Performance and carcass traits of goats fed on diets containing different proportions of mesquite (Prosopis chilensis (Molina) Stuntz.). Tropical Agriculture, 62(2):97-99.
JATASARA, D.S. and PARODA, R.S. (1981) Prosopis cineraria - an unexploited treasure of the Thar desert. Forage Research, 7(1):1-12.
JOSHI, U.N., ARORA, S.K., PARODA, R.S., JATASARA, D.S. and RANA, D.S. (1985) Chemical composition of Prosopis leaves to show the presence of anti-nutritional factors responsible for low in vitro dry matter digestibility. Nitrogen Fixing Tree Research Reports, 3:20-21.
KUMAR, R. (1983) Chemical and biochemical nature of fodder tree leaf tannins. Journal of Agricultural and Food Chemistry, 31(6):1364-1366.
LE HOUEROU, H.N. (1980) The role of browse in the Sahelian and Sudanian zones. In: Browse in Africa; The Current State of Knowledge. LE HOUEROU, H.N.(ed.) Addis Ababa, Ethiopia: International Livestock Centre for Africa. pp83-100.
LEAKEY, R.R.B. and LAST, F.T. (1980) Biology and potential of Prosopis sp. in arid environments, with particular reference to P. cineraria. Journal of Arid Environments, 3: 9-24.
LYON, C.K., GUMBMANN, M.R. and BECKER, R. (1988) Value of mesquite leaves as forage. Journal of the Science of Food and Agriculture, 44(2):111-117.
MARMILLON, E. (1986) Management of algarrobo (Prosopis alba, P. chilensis, P. flexuosa and P. nigra) in the semi-arid regions of Argentina. Forest Ecology and Management, 16(1/4):33-40.
McLEOD, M.N. (1974) Plant tannins: their role in forage quality. Nutrition Abstracts and Reviews, 44(11):803-815.
MENVIELLE, E.E. and HERNANDEZ, O.A. (1985) The nutritive value of calden pods (Prosopis caldenia Burk.). Revista Argentina de Produccion Animal, 5(7/8):435-439.
NAS (1979) Tropical Legumes: Resources for the Future. Washington D.C., USA: National Academy of Sciences.
PARTHASARATHY, M. (1986) Effect of feeding varying levels of Khejri (Prosopis cineraria) leaves and concentrate on the performance of weaner kids. Indian Journal of Animal Nutrition, 3(4):249-253.
PUNJ, M.L. (1988) Availability and utilization of non-conventional feed resources and their utilization by ruminants in South Asia. In: Non-conventional feed resources and fibrous agricultural residues: strategies for expanded utilization. DEVENDRA, C. (ed.). Proceedings of a Consultation held in Hisar, India, March, 1988. Ottawa, Canada: International Development Research Centre. pp50-61.
RAO, N.S.R. and REDDY, M.R. (1983) Utilization of Prosopis juliflora pods in the concentrate feeds of cattle and sheep. Indian Journal of Animal Sciences, 53(4):367-372.
ROBERTSON, A.G. (1980) Research being undertaken in the Atacama desert of Chile on the Tamarugo tree, Prosopis tamarugo Phil. In: Browse in Africa: The Current State of Knowledge. LE HOUEROU, H.N. (ed.). Addis Ababa, Ethiopia: International Livestock Centre for Africa. pp 389-391.
SEHGAL, J.P. (1984) Tannin content of different top feed leaves and utilization of tannin-rich pale (Ziziphus nummularia) leaves with concentrates for growing lambs. Indian Journal of Animal Sciences, 54(1):126-128.
SKERMAN, P.J., CAMERON, D.G. and RIVEROS, F. (1988) Tropical Forage Legumes. 2nd. edn. Rome: FAO. 692p.
TOUTAIN, B. (1980) The role of browse plants in animal production in the Sudanian zone of west Africa. In: Browse in Africa: The Current State of Knowledge. LE HOUEROU, H.N.(ed.) Addis Ababa, Ethiopia: International Livestock Centre for Africa. pp 103-108.
UPADHYAYA, R.S. (1985) Some nutritional and clinical observations in sheep fed khejri (Prosopis cineraria) tree leaves. Indian Journal of Animal Nutrition, 2(1):47-48.
UPHOF, J.C.T. (1968) Dictionary of Economic Plants. 2d edn. Lehre, Germany: J. Kramer. 591p.
VAITHIYANATHAN, S. and SINGH, M. (1989) Seasonal changes in tannin contents in some top feeds in arid region. Indian Journal of Animal Sciences, 59(12):1565-1567.
ZELADA, G.L. (1986) The �nfluence of the productivity of Prosopis tamarugo on livestock production in the Pampa del Tamarugal - a review. Forest Ecology and Management, 16(1/4):15-31.
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Use of Trees by Livestock (NRI, 1994, 160 p.)
Use of Trees by Livestock: Acacia
J.J. Bennison and R.T. Paterson
Acknowledgements
© Crown copyright 1993
The Natural Resources Institute (NRI) is an internationally recognized centre of expertise on the natural resources sector in developing countries. It forms an integral part of the British Government's overseas aid programme. Its principal aim is to alleviate poverty and hardship in developing countries by increasing the productivity of their renewable natural resources. NRI's main fields of expertise are resource assessment and farming systems, integrated pest management, food science and crop utilization.
NRI carries out research and surveys; develops pilot-scale plant, machinery and processes; identifies, prepares, manages and executes projects; provides advice and training; and publishes scientific and development material.
Short extracts of material from this publication may be reproduced in any non-advertising, nonprofit-making context provided that the source is acknowledged as follows:
Bennison, J.J., and Paterson, R.T. (1993) Use of Trees by Livestock 2: Acacia. Chatham, UK: Natural Resources Institute.
Permission for commercial reproduction should be sought from:
The Head, Publications and Publicity Section, Natural Resources Institute, Central Avenue, Chatham Maritime, Kent, ME4 4TB, United Kingdom.
No charge is made for single copies of this publication sent to the governmental and educational establishments, research institutions and non-profit-making organizations working in countries eligible for British Government Aid. Free copies cannot normally be addressed to individuals by name but only under their official titles.
Natural Resources Institute
ISBN: 0 85954 334-X
Price
£2.00LG2
This booklet has been printed on paper produced using the residue from sugar-cane processing.
Foreword
The importance of trees and shrubs in the feeding of animals in the tropics and sub-tropics has long been recognized by livestock owners. In arid areas where the growth of herbaceous plants is limited by lack of moisture, leaves and edible twigs of trees and shrubs can constitute well over 50% of the biomass production of rangeland. At high altitudes, tree foliage may provide over 50% of the feed available to ruminants in the dry season, branches being harvested and carried to the animals. Even in regions of higher rainfall where grass supplies the major proportion of the dry matter eaten by ruminants, tree leaves and fruits can form an important constituent of the diet, particularly for small ruminants.
In the last two decades interest in the planting of trees as a source of feed for livestock has been encouraged by workers in research and development, but in contrast to the hundreds of indigenous species which are used as fodder, attention has focussed on a limited number of introduced species. Thus there are many publications reporting the chemical composition of Leucaena leucocephala leaves and suggesting management strategies for utilization of the tree for fodder, but it is more difficult to find information on alternative genera which might be equally, or more, appropriate.
The aim of this series of publications is to bring together published information on selected genera of trees which have the potential to increase the supply of fodder for ruminants. Each booklet summarizes published information on the fodder characteristics and nutritive value of one genus, with recommendations on management strategies, where available. Further, since the leaves of woody species frequently contain secondary compounds which may have an anti nutritional, or toxic, effect, a separate booklet summarizes the effects of a number of these compounds. It is hoped that the booklets will provide useful resource material for students, research and extension workers, interested in promoting the use of trees as a source of fodder for ruminants.
Further copies of this booklet or others in the series can be obtained by writing to the Publications Section at the Natural Resources Institute.
Margaret Gill
Livestock Production
Programme
Genus Acacia
Family LEGUMINOSAE Subfamily MIMOSOIDEAE Tribe ACACIEAE
Principal phyllodenous species
Acacia aneura
Acacia combagei
Acacta cortacea
Acacia
cyanophylla
Acacia dealbata
Acacia decurrens
Acacia
holosericea
Acacia mearnsii
Acacia nilotica
Acacia pendula
Acacia
salicina
Acacia tortilis
Acacia trachycarpa (syn A. aff.
Iinarioides)
Acacia tumida
Principal bipinnate species
Faidherbia albida (syn. Acacia albida)
Acacia brevispica
Acacia cornigera
Acacia drepanolobium
Acacia farnesiana
Acacia
haematoxylon (syn. A. giraffae)
Acacia hebeclada subsp. hebeclada (syn. A.
stolonifera)
Acacia lasiopetala
Acacia leucophloea
Acacia mearnsii
(syn. Racosperma mearnsii)
Acacia mellifera
Acacia mellifera subsp.
detinens
Acacia nilotica
Acacia oerfota (syn. A. nubica)
Acacia
robusta
Acacia senegal
Acacia seyal
Acacia sieberiana
Acacia
tortilis
Acacia tortilis subsp. heteracantha (syn. A.
Iikatunensis)
Summary
With the African Acac�a spp. the fruit is probably more valuable as a feed than the foliage, because with the exception of F. albida, they tend to shed their leaves at the time of greatest feed scarcity during the dry season. Australian species however, retain their leaves and therefore show greater potential as fodder trees.
The productivity responses of animals reflect both the total intake, and the availability, of nutrients in the diet. Proximate and fibre analyses are unreliable indicators of nutritional value because of the presence of both cyanogenic glycosides and polyphenolics. A cautious approach should be adopted when feeding both A. leucophloea and some of the Australian species.
In some areas of Africa, Acacia spp. are the dominant browse trees within the natural ecosystem. Often there is no choice but to utilize the available feeds even if they do contain anti-nutritive compounds. Consumed as part of a wider diet, the effect of anti-nutritive factors is diluted. They only become serious in periods of extreme feed scarcity or in situations where the opportunity for feed selection is reduced, such as in stall feeding and cut-and-carry systems.
Acacia is a huge genus with pantropical distribution and a notable ability to survive in harsh environments. It has potential for use in environmental protection and for production of fodder, fuelwood, gums, and tannins.
Description and distribution
Acacia is a pantropical and subtropical genus with species abundant throughout Australia, Asia, Africa and the Americas. They thrive in a diverse range of habitats and environments. Many species are well adapted to the semi-arid and savannah regions but equally others survive in moist forest and riverine areas, tolerating both high pH and waterlogged soils. With such diversity, Acacia has considerable potential in a range of livestock and agroforestry systems. In Africa and Australia, some naturally occuring species are important in traditional pastoral and agropastoral systems, while imported species have become commercially accepted. Trees provide fodder and shade for livestock, improve soil fertility through nitrogen fixation and the production of leaf litter end stabilize veils. Acacia species provide edible fruits and seeds, gum arabic and timber for fuel, construction and fencing.
In a large genus of 800-900 species (Allen and Allen, 1981), classification and nomenclature can be complex, particularly as there have been a number of revisions and attempts to subdivide the genus. Pedley (1986; 1987) advocated that it be divided into three separate genera, namely Acacia, with 200 species best represented in Africa and South America, Senegalia, with about 150 species with the same geographical distribution, and Racosperma, with about 850 species virtually confined to Australia. While the suggestion may have merit, it has not yet been generally accepted. This text uses the traditional nomenclature adopted by standard flora. Where a name has been changed as a result of a generally accepted revision, the name used in the original reference is listed as a synonym. The names of the African species are those included in the check-list produced by Lock (1989) as part of the programme of the International Legume Database and Information Service (ILDIS).
One widely accepted recent change is the reclassification of Acacia albida as Faidherbia albida, although Coe and Beentje (1991) stated that the justification for the change was primarily phytochemical. In this publication, for purposes of discussion and comparison, the name F. albida is accepted but it is considered as though it had remained within the genus Acacia, since it shares many agronomic characteristics with species that have not been reclassified.
Various workers have divided the genus into divisions, series and subseries (Bentham, 1864) or sections (Taubert, 1894) but for ease of description and an understanding of the fodder potential, it is simplest to describe Acacia spp. as being either phyllodenous (principally Australian in origin) or bipinnate (mainly from Africa and Asia).
Phyllodenous Acacia species
The majority of Australian Acacia spp. lack true leaves. Instead, they exhibit a characteristic known as phyllodony where the petiole is expanded and flattened. The phyllode has the ability to photosynthesize and for all practical purposes, it is the equivalent of a leaf. The Australian exceptions are from the Section Botrycephalae, which maintain bipinnate foliage until maturity and do not develop phyllodes.
Vercoe (1987) noted that species of Acacia and other genera from the dry tropics of Australia had potential in the semi-arid zones of Africa for fuelwood and erosion control. If their usefulness as fodder could also be established, this would enhance their overall value. Certainly the value of trees such as A. aneura as drought reserve feed is well established and recognized by pastoralists in Australia. Unfortunately, success in establishing A. aneura in Africa has been limited. However, a number of other species appear to show potential, notably A. holosericea, A. trachycarpa and A. coriacea. Other species that have been studied include A. salicina and A. cyanophylla, although the phyllodes of the latter contain high tannin levels (Vercoe, 1987; Goodchild and McMeniman, 1987; Reed et al., 1990), which can reduce both the palatability and the feeding value. Unlike African and Asian species, Australian Acacia spp. are evergreen and unarmed.
Bipinnate Acacia species
There are about 200 bipinnate Acacia species in the New World and a further 150 species in Africa and Asia. All 129 African species possess bipinnate leaves. There are other physical and physiological characteristics which separate them from their Australian relatives, the Botrycephalae. This group, the best known of which is A. mearnsii (black wattle), grown throughout the subtropical world for the extraction of tannin, is confined to the cooler and moister parts of south eastern Australia. They are not noted as valuable fodder trees (Boland, 1987). Most bipinnate Acacia spp. are armed with thorns of various shapes and sizes, some of which are modified stipules at the base of the leaves. Many exhibit transient periods of defoliation in the dry season and have been described as deciduous. Before the start of the rains, there is a often a flush of new growth of leaves. Although the stimulus for this activity is uncertain, it may be due to temperature.
Fodder characteristics
Fodder yield
Information on leaf yield of African bipinnate Acacia spp. is limited, and comparative data with introduced Australian species, even more so. The work of Cossalter (1986) compared species from the two regions, when grown in Senegal at uniform spacings of 3 x 3 m (Table 1). The results, as means of 20, or 25 individual trees aged between 18-23 months, demonstrated the fodder potential of the Australian Acacia species when compared with African ones. A. holosericea is perhaps the most frequently planted Australian Acacia in development projects because of its superior yield and because it retains a large phyllode biomass during the dry season, while African species shed their leaves during this period (Cossalter, 1986). There is some uncertainty about the palatability of A. holosericea: early trials indicate that only dry phyllodes are accepted. The fresh phyllodes of A. trachycarpa and A. tumida, however, were readily eaten by both cattle and sheep (Hamel, 1980).
Table 1 - Total biomass production (phyllodes, leaves and wood) of Acacia spp.
|
Origin phyllodes |
Species(kg/tree) |
Dry leaves, (g/tree) |
Wood |
|
Australia |
A. Holosericea |
2660 |
12.03 |
|
Australia |
A. trachycarpa |
1993 |
10.65 |
|
Australia |
A. tumida |
2600 |
8.69 |
|
Australia |
A. trachycarpa |
1700 |
3.73 |
|
Africa |
A. senegal |
60 |
9.92 |
|
Africa |
A. seyal |
208 |
9.39 |
|
Africa |
A. tortilis |
130 |
5.89 |
Source: Cossalter (1986)
When assessing the fodder value of African Acacia spp., the importance of the fruit and pods cannot be ignored. This fraction may be of more value than the foliage in some cases. A major problem in estimating the yield of fruit is the variation in individual trees between years. Trees may bear a heavy crop one year and little the next. The fruits of F. albida are relished by game, cattle and small ruminants and NAS (1979) suggests that yields of 125-135 kg are possible from a single tree. When ripe, the fruit can be harvested and stored for future use, although there is a tendency for the pods to shatter and disintegrate.
The bark of A. seyal is particularly valuable as a fodder, although complete removal will result in the death of the tree. Skerman et al., (1988) suggested that the bark, which is extensively used for feeding cattle and small ruminants in the dry season, was the most valuable part of the plant. Cattle could consume up to 5.5 kg bark/day, sufficient for maintenance and the production of 4.5 litres of milk.

Table 2 - Mean proximate and fibre
analyses of Acacia leaves, phyllodes, fruits and
seeds
Nutritive value
In Table 2, the mean proximate and fibre analyses of leaves, phyllodes and fruits of Acacia spp. are summarized from data of individual species (Appendix 1). Absolute values should be treated with caution since there is considerable variation in the literature. For example, the mean crude protein (CP) level of Acacia leaves is 18.9% of dry matter (DM), with a reported range of 6.5-42.8%. However, the data provide an indication of the nutritive value and in general both leaves and fruits of all species are valuable sources of protein, particularly when compared with mature grasses, where CP levels in forage dry matter (DM) during the dry season can fall to 3-4% or less, well below the 7% CP which is suggested as a lower limit for fibre digestion in cattle (Van Soest, 1982).
While the crude fibre (CF) levels of Acacia are relatively high (15-40%) compared with other fodder tree species such as Leucaena leucocephala and Gliricidia septum, they are still markedly lower than for tropical grasses and common crop residues. Even the fruit, consisting of the dry dehiscent pod and the seeds contained in it, are a valuable feed source, with moderate CF (25%) and high CP levels (15%). The phyllodenous Acacia spp. tend to have marginally higher CF and lower CP levels than bipinnate species (Appendix 2) though the available data consist of fewer observations. The lower nutritive value is offset by both the higher biomass yield and the retention of the phyllodes throughout the dry season, when bipinnate species tend to shed their leaves.
Information on the digestibility of DM and organic matter (OM) of Acacia leaves and phyllodes is limited but the available data indicate that, with sheep, they are relatively low and in the range of 40-605 to respectively (Table 3). The digestibility of phyllodes appears to be lower than that of bipinnate leaves. Goodchild and McMeniman, (1987) suggested that the low fibre digestibility could be associated with the high lignin content of the cell wall, since fibre digestibility is inversely related to the lignin content of the fibre fraction.
Information on the digestibility of DM and organic matter (OM) of Acacia leaves and phyllodes is limited but the available data indicate that, with sheep, they are relatively low and in the range of 40-60% respectively (Table 3). The digestibility of phyllodes appears to be lower than that of bipinnate leaves. Goodchild and McMeniman, (1987) suggested that the low fibre digestibility could be associated with the high lignin content of the cell wall, since fibre digestibility is inversely related to the lignin content of the fibre fraction.
While the digestibility of the whole fruit (seed and pod) is higher than that of the leaves and phyllodes, it can be adversely affected by the proportion of seeds that pass undigested through the gut of the animal. Tanner et al., (1990) reported that 46% of A. tortilis seeds were voided in the faeces of sheep. Similarly, Goodchild and McMeniman (1987) noted that when feeding yearling cattle with seeds of A. tortilis subsp. spirocarpa, 12% passed unchanged through the gut. Legume seeds are physically hard and notoriously resistant to digestive juices, although the small size of Acacia seed may be a contributory factor in their passage through the gut (Tanner et al., 1990).
The digestibility of the leaf and fruit fraction can also be depressed by the presence of polyphenolic compounds, including tannins.
Anti-nutritive factors
Acacia spp. have acquired a variety of physical and chemical defence mechanisms aimed at reducing the palatability and nutritive value of the plant to predators including both insects and animals.

Table 3 - In vivo
digestibility (%) of Acacia forage
Perhaps the most obvious physical characteristic is the spiny armature of the African species, which evolved as a defence against large herbivores. A. mellifera subsp. detinens derived its common name (wait-a-bit tree) from the pairs of small hooks on the branches. A. tortilis is protected by two types of thorns, long white stipular spines and small curved hooks, a formidable protection against animals. Coe and Beentje (1991) suggested that as thorns restrict the availability of forage, a critical level is reached below which the herbivore no longer finds it nutritionally efficient to seek shoots between the spines, since this invariably involves movement between each bite. There is also evidence that in A. drepanolobium, thorns on branches which are regularly browsed by animals are significantly longer than those on undefoliated branches (Young, 1987).
Tree size is a form of physical protection that can be harnessed as a desirable characteristic in certain agroforestry systems. Domestic animals are unable to browse tall trees which then maintain a standing reserve of foliage which can be made available by strategic lopping at critical periods when other feed is scarce.
A fascinating defence mechanism of the genus is the symbiotic relationship with ants known as myrmecophily, where the plant lives in association with an ant colony. In the tropical American species A. cornigera, association with ants from the genus Pseudomyrmex is a notable example of myrmecophily. The trees have modified, swollen spines or galls, which provide shelter for the ants, and extrafloral nectaries and modified leaflet tips which provide food. The ants, in return, protect the trees from insects and other predators by swarming to attack them when the tree is disturbed (Janzen, 1966). In Africa, A. drepanolobium is commonly known as the 'whistling thorn', a name derived from the noise of the wind blowing into the holes of the galls that have been occupied by ants.
Many Acacia spp. produce a range of potentially toxic compounds which prevent herbivores from eating the foliage, although it is thought that the chemicals originally evolved as defence mechanisms against insect and fungal attack. Some members contain specific anti-nutritive factors, for example, A. georginae has been implicated in heavy stock losses in Australia due to its content of fluoracetic acid (D'Mello, 1992). In general terms the genus contains two major groups of anti-nutritive factors, cyanogenic glycosides and polyphenolic compounds.
Cyanogenic glycosides
When cyanogenic substrates are broken down under the action of a hydrolyzing enzyme, hydrogen cyanide (HCN) is formed. Plants that contain both cyanogenic glycosides and an endogenous, hydrolytic enzyme are the most dangerous but broad-spectrum enzymes may be present in other fractions of the diet. Symptoms of HCN poisoning include respiratory dyspnoea, intense red conjunctive, frothing at the mouth, bloat, a staggering gait, convulsions and violent death. On post-mortem examination, a characteristic almond smell is often noted in the contents of the gut.
In Asia, there are a number of reports of HCN poisoning of livestock when fed on the pods and leaves of A. Ieucophloea (Gupta and Naurival, 1966; Prasad et al., 1977; Vihan and Panwar, 1987). In South Africa, Steyn and Rimington (1935) reported the presence of cyanogenic glycosides in A. haematoxilon, A. Iasiopetala, A. tortilis subsp. heteracantha, A. robusta and A. hebeclada subsp. hebeclada. They found higher concentrations of HCN from leaves and immature pods than from mature pods. In A. robusta, the glycoside was present without the corresponding hydrolytic enzyme. They isolated the cyanogenic compound acacipetalin from A. Iasiopetala and A. hebeclada. Siegler et al., (1975) isolated a further compound, dihydroacipetalin, from the leaves and shoots of A. sieberiana var. woodii.
Toxicity appears only in sporadic outbreaks, being dependent on a number of factors, including the species and variety of Acacia, the rate of ingestion by the animal and the availability of alternative feeds. It is more likely to occur during periods of drought or feed scarcity when hungry animals consume large quantities of feed in a short period of time. Access to cold water during feeding appears to promote the release of HCN from the substrates. Practical measures can be taken to reduce the risk of poisoning such as restricting the intake, particularly of leaves and immature pods, separating the time or place of feeding and watering, and feeding dangerous material with molasses or sulphur. These substances combine with HCN to form harmless cyanhydrin and sulphocyanides respectively (Steyn and Rimington, 1935).
Phenolic compounds
There are numerous reports of the presence of polyphenolic compounds in the various components of plants of the genus Acacia. From the standpoint of nutrition, the most important of these are the tannins, compounds with molecular weights in excess of 500, which are able to form complexes with, and to precipitate proteins (McLeod, 1974; Mangan, 1988). Hydrolyzable tannins are readily broken down by acids, bases and enzymes to form sugars and phenolic carboxylic acid and are therefore easily degraded in the gut. In contrast, condensed tannins are not degraded and so are available to form complexes with proteins, resulting in reduced feed intake and digestibility. McNaughton (1987) suggested that at levels above about 5%, increasing the tannin content in a diet reduced its palatability.
Reed et al., (1990) assessed the effects of polyphenolics on intake, growth, digestibility and nitrogen utilization by sheep. Three Acacia forages, (phyllodes of A. cyanophylla, leaves of A. seyal, and fruit of A. sieberiana), all containing high levels of polyphenolics, were compared with three protein supplements and another fodder tree Sesbania sesban which is low in polyphenolics. All diets were fed in combination with teff straw (Eragrostis abyssinica). The diets containing Acacia led to the highest levels of feed refusal by the sheep their and lowest intake of teff straw. Fibre digestibility was lower for these diets and the tannins complexed with proteins, resulting in higher levels of faecal nitrogen.
A complementary study by Tanner et al., (1990) reported significant differences in the growth rates of sheep supplemented with fruits from four different Acacia spp. (A. tortilis, F. albida, A. nilotica and A. sieberiana) even though there were no significant differences in total nitrogen intake. The tannin-rich fruits, particularly of A. sieberiana and A. nilotica, reduced feed intake and the efficiency of nitrogen utilization by forming complexes with proteins which again resulted in a shift of nitrogen excretion from urine to faeces. The growth rates of sheep fed the diets low in polyphenolics (F. albida and A. tortilis), were not significantly different from sheep supplemented with noug meal (Guizotia abyssinica), indicating their suitability as dietary supplements.
The literature abounds with conflicting reports on the suitability of Acacia spp. for livestock feed.
Dumancic and Le Houerou (1981) considered the phyllodes of A. cyanoophylla to be sufficient to meet the protein maintenance requirements of sheep, while Reed et al., (1990) reported a negative nitrogen balance for animals fed on this legume. Predictions of the productive responses of livestock are problematical and uncertain. There are a number of possible explanations for the conflicting reports:
· the chemistry of tannins is complex and analytical results do not always correlate with biological effects, e.g. nitrogen digestibility;
· a single feed may contain more than one antinutritive factor. A. nilotica is known to have both tannins and catechin gallates (Mueller-Harvey et al., 1987);
· the concentration and type of phenolic compound both vary with season and in response to stimuli such as defoliation;
· there is much variation between both species and provenance in tannin content, the effect being both genetic and environmental.
Management
Where the principal use of Acacia spp. is for browsing by livestock, little is done to manage the resource, except through some measure of defoliation control by herding the animals. High branches are lopped during times of feed shortage, particularly in the dry season. At other times of the year, the trees are subjected to uncontrolled browsing, often by both wild and domestic animals.
Many legumes produce seed which has a hard, impermeable seed coat. Such seed will not germinate until moisture is able to penetrate it. This characteristic, known as 'hard seededness', prevents the simultaneous germination of the entire stock of seed in the soil at the start of the wet season. This adaptation favours the long-term survival of the population where rainfall is unpredictable. It is, however, a disadvantage when attempting to establish new stands from seed, particularly in areas with a short growing season. Acacia shows extreme variability in the degree of hard seededness, both between and within species; the proportion of hard seed in a sample depends on the environmental conditions during plant growth, the maturity of the seed at harvest and the length of time it has spent in storage. In general, Australian species respond to seed scarification. Recommended treatments (Doran and Gunn, 1987) include:
· manual nicking of the seed coat. Because of the amount of labour involved, this is suitable only for seed for germination tests, or for valuable research lots;
· placing the seed in boiling water for one minute. This is recommended for many hard-coated seed lots;
· placing the seed in hot water (80-90°C) for one minute. This is recommended where sensitivity to boiling water is suspected.
In Central Australia where annual rainfall is about 300 mm, native Acacia spp. naturally regenerate after exceptionally high rainfall at populations of some 5000 stems/ha, and many species produce multiple stems. Subsequent fires and drought will thin the stand to 100-400 stems/ha. Under these conditions,
the optimum stocking level for planted stands may be in the region of 1000 seedlings/ha, followed by thinning as the stand matures. Alternatively, where thinning is not practical, an initial stand of about 400 trees/ha would be appropriate (Kube, 1987). The seed is scarified in hot water (80°C) and germinated in trays. Seedlings are transplanted after a few days into individual tubes 5 cm in diameter by 15 cm deep, filled with a potting mixture of equal parts of coarse sand, fine sand or sandy loam and peat moss. Treatment with fungicide is usually necessary and the seedlings are grown in decreasing shade for about three months before planting out. Supplementary irrigation is applied for 6-12 months, depending upon rainfall. Growth is often rapid, many species attaining heights of 2 m in about two years.
In Tanzania, A. mearnsii plantations have been established for fuelwood, charcoal and building poles by direct seeding scarified seed at rates of 1.22.4 kg/ha in rows 1.8 m apart in fully cultivated soil (Kessy, 1987). The major problem with this species has been its susceptibility to competition from weeds. Harvesting has been on an 8-12 year cycle and natural regeneration has taken place after the original stand has been clear felled and burned.
Alternative uses
While Acacia is the largest of the mimosoid genera and the second largest of all the legume family, only about 75 members have economic value, and of these only about 50 are actually cultivated. Nevertheless, they are used in various ways, as described by Allen and Allen, (1981) and Boland, (1987).
Many Acacia spp. form spiny, slow-growing scrub which provides protection against soil erosion caused by wind and rain. Their tolerance of harsh conditions makes them valuable for stabilization and revegetation of difficult sites. A. armata, A. glaucescens, A. pycnantha and A. suaveolens tolerate salt spray and so can be used to stabilize coastal sand dunes. A. decurrens is planted along roadsides in Australia and on denuded hills in Rwanda. A. mearnsii and A. dealbata are used to control gulley and hillside erosion in New Zealand.
Acacia is an important source of fuelwood or charcoal in arid regions. A. mearnsii is used for both domestic fuel and tobacco curing in Indonesia (NAS, 1980). It also provides poles for mining and agriculture in many parts of the world.
As timber, Acacia wood is extremely durable and although it is hard to work, it finishes well. Smaller species provide wood which is used to make implement handles, pipes and furniture, while larger species are used for paneling, boats and musical instruments. Hawaian mahogany (A. koa) was once prized for making surfboards, canoes and bowls while A. segal and A. tortilis were the principal trees of the Arabian Desert region. A. glaucescens and A. melanoxylon are major timber trees today. A. dealbata and A. mearnsii are used for pulping in Australia and Japan, while the latter species is used in the production of rayon in South Africa and India.
A. mearnsii is the world's most important source of vegetable tannin, 35-40% of the air-dry weight of bark is extracted for leather manufacture. Yields in excess of 15% are also obtainable from other species such as A. albida and A. cyanophylla. Wood adhesives and polyurethane foams can be prepared from bark extracts of A. mearnsii, and used as substitutes for petroleum products.
Gum arable, an important emulsifier, protective colloid, adhesive and binder, is produced in commercially exploitable quantities by A. arabica, the principal historical source, and more recently by A. senegal and some 20 other species of Acacia. It is used in foods, pharmaceuticals, cosmetics, adhesives, paints, polishes and inks and in lithography, photography, textile-sizing and other processes.
Species such as A. dealbata, known as mimosa by florists, A. albida and A. tortilis, are valued as ornamentals while A. farnesiana and A. dealbata are used in the production of base oils for perfumes. In Australia, several species are valuable sources of pollen in apiculture.
References and further reading
AHN, J.H., ROBERTSON, B.M., ELLIOT, R., GUTTERIDGE, R.C. and FORD, C.W. (1989) Quality assessment of tropical browse legumes: tannin content and protein degradation. Animal Feed Science and Technology, 27: 147-156.
ALLEN, O.N. and ALLEN, E.K. (1981) The Leguminosoe: a Source Book of Characteristics, Uses and Nodulation. Madison, USA: University of Wisconsin Press.
BATE-SMITH, E.C. (1973) Haemanalyis of Tannins: The Concept of Relative Astringency. Phytochemistry, 12: 907-912.
BENTHAM, G. (1864) Flora Australiensis vol.2. London: Reeve and Co.
BOLAND, D. J. (1987) Genetic resources and utilisation of Australian bipinnate acacias (Botrycephalae). In: Australian Acacias in Developing Countries. TURNBULL, J.W. (ed.) Proceedings, International Workshop held at Gympie, Australia, August, 1986. ACIAR Proceedings No.16. pp. 29-37.
BUTLER, L.G. (1982) Relative degree of polymerization of sorghum tannin during seed development and maturation. Journal of Agriculture and Food Chemistry, 30: 1090-1094.
COE, M. and BEENTJE, H. (1991) A Field Guide to the Acacias of Kenya. Oxford, UK: Oxford University Press.
COSSALTER, C (1986) Introduction of Australian Acacias into Dry Tropical West Africa. Forest Ecology and Management, 16: 367-389.
D'MELLO, J.P.F. (1992) Chemical Constraints to the use of tropical legumes in animal nutrition. An�mal Feed Science and Technology, 38: 237-261
DORAN, J.C. and GUNN, B.V. (1987) Treatments to promote seed germination in Australian Acacias. In: Australian Acacias in Developing countries. TURNBULL, J.W. (ed.) Proceedings, International Workshop held at
Gympie, Australia, August 1986. ACIAR Proceedings No. 16. pp. 57-63.
DUMANCIC, D. and LE HOUEROU, H.N. (1981) Acacia cyanophylla Lindl. as a supplementary feed for small stock in Libya. Journal of Arid Environments, 4: 161 -167.
GOHL, B. (1981) Tropical Feeds: Feed Information Summaries and Nutritive Values. FAO Animal Production and Health Series No. 12. Rome: FAO.
GOODCHILD, A.V. and McMENIMAN, N.P. (1987) Nutritive value of Acacia foliage and pods for animal feeding. In: Australian Acacias in Developing Countries. TURNBULL, J.W. (ed.) Proceedings, International Workshop held at Gympie, Australia, August 1986. ACIAR Proceedings No.16. pp. 101-106.
GUPTA, I. and NAURIYAL, M.M. (1966) Acacia leucophloea Willd (raunja) poisoning in livestock. Indian Veterinary Journal, 43: 538-543.
HAGERMAN, A.E. and BUTLER, L.G. (1981) The specificity of proanthocyanidin-protein interactions. Journal of Biological Chemistry, 256: 4494-4497.
HAMEL, O. (1980) Acclimatization and utilization of phyllodineous acacias from Australia in Senegal. In: Browse in Africa; The current state of knowledge. LE HOUEROU, H.N. (ed.) International Livestock Centre for Africa: Addis Ababa. pp. 361-373.
JANZEN, D.H. (1966) Coevolution of mutualism between ants and acacias. Evolution, 20: 249-275.
KESSY, B.S. (1987) Growth of Australian Acacias in Tanzania. In: Australian Acacias in Developing Countries. TURNBULL, J.W. (ed.) Proceedings, International Workshop held at Gympie, Australia, August 1986. ACIAR Proceedings No. 16. pp. 123-125.
KUBE, P.D. (1987) Growth rates, establishment techniques and propagation of some Central Australian Acacias. In: Australian Acacias in Developing Countries. TURNBULL, J.W. (ed.) Proceedings, International Workshop held at Gympie, Australia, August 1986. ACIAR Proceedings No. 16. pp. 77-80.
KUMAR, R. and VAITHIYANATHAN, S. (1990) Occurrence, nutritional significance and effect on animal productivity of tannins in tree leaves. Animal Feed Science and Technology, 30: 21-38.
LE HOUEROU, H.N. (1980) The role of browse in the Sahelian and Sudanian zones. In: Browse in Africa; The current state of knowledge. LE HOUEROU, H.N. (ed.) International Livestock Centre for Africa: Addis Ababa. pp. 83-100.
LOCK, J.M.(1989) LegumesofAfrica: ACheckList. Kew: Royal Botanic Gardens.
MAKKAR, H.P.S., DAWRA, R.K. and SINGH, B. (1988) Changes in tannin content, polymerisation and protein precipitation capacity in oak (Quercus incana) leaves with maturity. Journal of Science of Food and Agriculture, 44: 301-307.
MANGAN, J.L. (1988) Nutritional effects of tannins in animal feeds. Nutrition Research Reviews, 1: 209-231.
MASLIN, B.R., CONN, E.E. and DUNN, J.E. (1987) Cyanogenic Australian species of Acacia: A preliminary account of their toxicity potential. In: Australian Acacias in Developing Countries. TURNBULL, J.W. (ed.) Proceedings, International Workshop held at Gympie, Australia, August 1986. ACIAR Proceedings No.16. pp. 107-111.
McLEOD, M.N. (1974) Plant tannins - their role in forage quality. Nutrition Abstracts and Reviews, 44: 803-815.
McNAUGHTON, S.J. (1987) Adaption of herbivores to season changes in nutrient supply. In: Nutrition of Herbivores. HACKER, J.B. and TERNMOUTH, J.H. (eds). Sydney, Australia: Academic Press. pp. 391-408.
MUELLER-HARVEY, I., REED, J.D. and HARTLEY, R.D. (1987) Characterisation of phenolic compounds, including flavonoids and tannins, of ten Ethiopian browse species by high performance liquid chromatography. Journal of the Science of Food and Agriculture, 39: 1-14.
NAS (1979) Tropical Legumes: Resources for the Future. Washington DC: National Academy of Sciences.
NAS (1980) Firewood Crops. Washington DC: National Academy of Sciences.
PEDLEY, L. (1986) Deprivation and dispersal of Acacia (Leguminosue) with particular reference to Australia, and recognizing Senegalia and Racospenna. Botanical Journal of the Linnean Society, 92: 219-54.
PEDLEY, L. (1987) Australian Acacias: Taxonomy and Phytogeography. In: Australian Acacias in Developing Countries. TURNBULL, J.W.(ed.) Proceedings, International Workshop held at Gympie, Australia, August 1986. ACIAR Proceedings No 16. pp. 11-16.
PRASAD, J., SINGH, A.P. and REKIB, A. (1977) Hydrocyanic acid poisoning in grazing sheep and goat on Acacia leucophloca (reunja). Indian Veterinary Journal, 54: 748-751.
REED, J.D., HORVARTH, P.J., ALLEN M.S. and VAN SOEST, P.J. (1985) Gravimetric determination of soluble phenolics including tannins from leaves byprecipitation with trivalent ytterbium. Journal of the Science of Food and Agriculture, 36: 255-261.
REED, J.D., SOILER, H., and WOODWARD, A. (1990). Fodder tree and straw diets for sheep: intake, growth, digestibility and the effects of phenolics on nitrogen utilisation. Animal Feed Science and Technology, 30: 3950.
SEDDON, H.R. and KING, R.O.C. (1930) As to the fatal dose for sheep of cyanogenetic plants containing Sambunigrin and Prunasin. Journal Council Scientific and Industrial Research Australia, 3: 14-24.
SIEGLER, D.S, BUTTERFIELD, C.S., DUNN, J.E. and CONN, E.E. (1975). Dihydroacacipetalin - a new cyanogenic glycoside from Acacia sieberiana var woodii. Phytochemistry, 14: 1419-1420.
SKERMAN, P.J., CAMERON, D.G. and RIVEROS, F. (1988) Tropical Forage Legumes. 2nd. edn. Rome: FAO.
STEYN, D.G. and RIMINGTON, C. (1935). The occurrence of cyanogenetic glucosides in South
African species of Acacia. Onderstepoort Journal of Veterinary Science and Animal Industry, 4(1): 51-73.
TANNER, J.C., REED, J.D. and OWEN, E. (1990). The nutritive value of fruits (pods with seeds) from four Acacia spp. compared with extracted noug (Guizotia abyssinica) meal as supplements to maize stover for Ethiopian highland sheep. Animal Production, 51: 127-133.
TAUBERT, P. (1894) Die Naturlichen Planzenfamilie. Leipzig: Wilhelm Engelmann.
TERBLANCE, M., PIENAAR, J.G., BIGALKE, R. and VAHRMEYER, J. (1967) Acacia nilotica (L.) Del. subsp. kraussiana (Berth) Brenan. as a poisonous plant in South Africa. Journal South African Veterinary Medical Association, 38(1): 57-63.
VAN HOVEN, W. (1985) The tree's secret weapon. South African Panorama, 30(3): 34-37.
VAN SOEST, P.J. (1982) Nutritional Ecology of the Ruminant. Corvallis, Oregon, USA: O & B Books.
VERCOE, T.K. (1987) Fodder potential of selected Australian tree species. In: Australian Acacias in Developing Countries. TURNBULL, J.W. (ed.) Proceedings, International Workshop held at Gympie, Australia, August 1986. ACIAR Proceedings No.16. pp. 95-100.
VIHAN, V.S. and PANWAR, H.S. (1987) A note on toxicity of Acacia leucophloea (Rajma) in sheep. Indian Journal Animal Research, 21(1): 53-55.
WILSON, A.D. (1969). A review of browse in the nutrition of grazing animals. Journal of Range Management, 22: 23-28.
WOODWARD, A. and REED, J.D. (1989) The influence of polyphenolics on the nutritive value of browse: A summary of research conducted at ILCA. ILCA Bulletin 35, Addis Ababa: International Livestock Centre for Africa.
YOUNG, T.P. (1987) Increased thorn length in Acacia drepanolobium - an induced response to browsing. Oecologia 71(3): 436-438.
Appendix 1
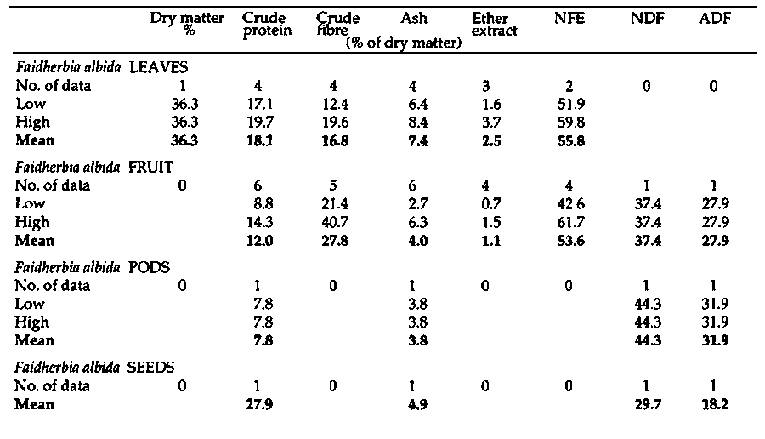
Primate and fibre analyses of
bipinnate Acacia spp.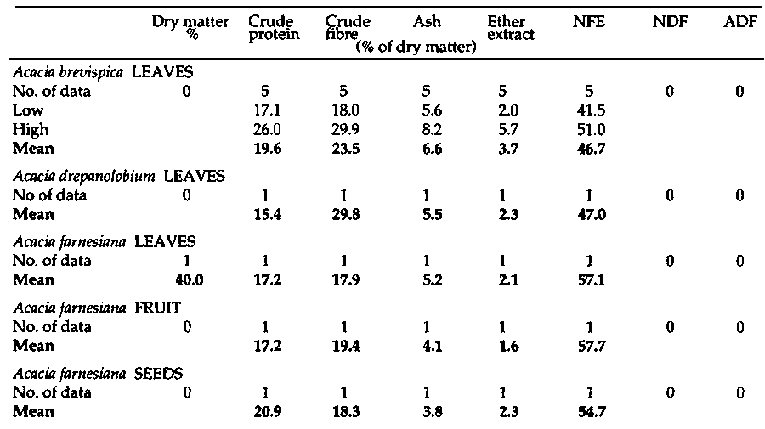
Primate and fibre
analyses of bipinnate Acacia spp. (continued 1)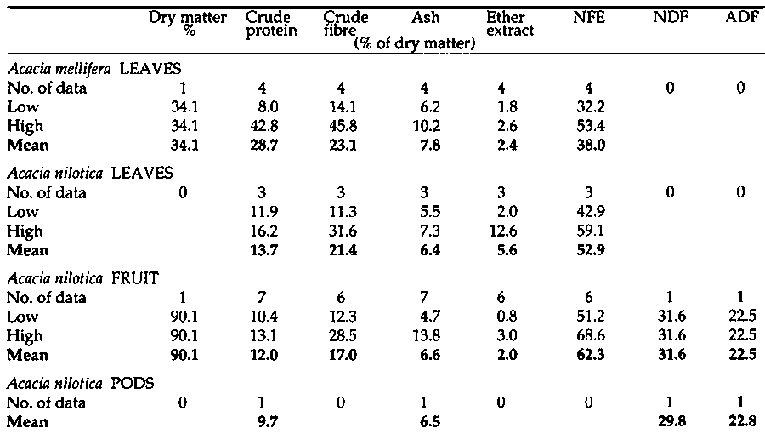
Primate and fibre analyses of bipinnate Acacia spp. (continued
2)
Primate and fibre analyses of bipinnate
Acacia spp. (continued 3)
Primate and fibre
analyses of bipinnate Acacia spp. (continued 4)
Primate and fibre analyses of bipinnate Acacia spp. (continued
5)
Primate and fibre analyses of bipinnate
Acacia spp. (continued
6)
Appendix 2

Proximate analyses of Acacia
phyllodes
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Use of Trees by Livestock (NRI, 1994, 160 p.)
Use of Trees by Livestock: Gliricidia
J.J. Bennison and R.T. Paterson
© Crown copyright 1993
The Natural Resources Institute (NRI) is an internationally recognized centre of expertise on the natural resources sector in developing countries. It forms an integral part of the British Government's overseas aid programme. Its principal aim is to alleviate poverty and hardship in developing countries by increasing the productivity of their renewable natural resources. NRI's main fields of expertise are resource assessment and farming systems, integrated pest management, food science and crop utilization.
Acknowledgement
NRI carries out research and surveys; develops pilot-scale plant, machinery and processes; identifies, prepares, manages and executes projects; provides advice and training; and publishes scientific and development material.
Short extracts of material from this publication may be reproduced in any non-advertising, nonprofit-making context provided that the source is acknowledged as follows:
Bennison, J.J., and Paterson, R.T. (1993) Use of Trees by Livestock 3: Gliricid ia. Chatham, UK: Natural Resources Institute.
Permission for commercial reproduction should be sought from:
The Head, Publications and Publicity Section, Natural Resources Institute, Central Avenue, Chatham Maritime, Kent, ME4 4TB, United Kingdom.
No charge is made for single copies of this publication sent to the governmental and educational establishments, research institutions and non-profitmaking organizations working in countries eligible for British Government Aid. Free copies cannot normally be addressed to individuals by name but only under their official titles.
Natural Resources Institute ISBN: 0 85954 3331 Price £2.00 LG3
This booklet is printed on paper produced using the residues from sugar-cane processing.
Foreword
The importance of trees and shrubs in the feeding of animals in the tropics and sub-tropics has long been recognized by livestock owners. In arid areas where the growth of herbaceous plants is limited by lack of moisture, leaves and edible twigs of trees and shrubs can constitute well over 50% of the biomass production of rangeland. At high altitudes, tree foliage may provide over 50% of the feed available to ruminants in the dry season, branches being harvested and carried to the animals. Even in regions of higher rainfall where grass supplies the major proportion of the dry matter eaten by ruminants, tree leaves and fruits can form an important constituent of the diet, particularly for small ruminants.
In the last two decades interest in the planting of trees as a source of feed for livestock has been encouraged by workers in research and development, but in contrast to the hundreds of indigenous species which are used as fodder, attention has focussed on a limited number of introduced species. Thus there are many publications reporting the chemical composition of Leucanena leucocephala leaves and suggesting management strategies for utilization of the tree for fodder, but it is more difficult to find information on alternative genera which might be equally, or more, appropriate.
The aim of this series of publications is to bring together published information on selected genera of trees which have the potential to increase the supply of fodder for ruminants. Each booklet summarizes published information on the fodder characteristics and nutritive value of one genus, with recommendations on management strategies, where available. Further, since the leaves of woody species frequently contain secondary compounds which may have an antinutritional, or toxic, effect, a separate booklet summarizes the effects of a number of these compounds. It is hoped that the booklets will provide useful resource material for students, research and extension workers, interested in promoting the use of trees as a source of fodder for ruminants
Further copies of this booklet or others in the series can be obtained by writing to the Publications Section at the Natural Resources Institute.
Margaret Gill
Livestock Production
Programme
Genus Gliricidia
Family LEGUMINOSAE
Subfamily PAPILIONOIDEAE
Tribe
GALEGEAE
Subtribe ROBINIINAE
Principal species Gliricidia
ehrenbergii
Gliricidia maculata
Gliricidia septum
Main common names
Quickstick (Caribbean)
Mata Raton (Puerto Rico)
Madre Cacao (Central
America)
Summary
Gliricidia has long been used for shade and physical support for a range of crops and as living fence posts. It has toxic effects on rodents and possibly horses. It can be a useful supplement for ruminants, even though palatability appears to be a problem with some accessions under certain conditions. The major drawback in many systems is its tendency to shed its leaves during the dry season, although this effect can be minimized by frequent cutting during the wet season, together with storage of surplus foliage as either sun-dried leaf meal, or as silage prepared with molasses and formic acid as additives. Presently exploited accessions represent only a small part of the existing germplasm held in international collections and there is much scope for selection within existing gene banks. There may also be scope for further collection from native populations before it becomes necessary to resort to breeding to achieve further improvements of exploitable characteristics. The tree shows potential for tropical animal production systems on relatively acid, infertile soils, particularly in sub-humid and humid regions and is deserving of further evaluation and development.
Description and distribution
Gliricidia is a genus of six to nine species of small, spreading, unarmed, fast-growing perennial shrubs or short-bored trees, reaching 5-15 m in height. They are able to fix nitrogen and nodulation has been observed and evaluated (Patil and Prasunamma, 1986). They nodulate readily, usually within three months of planting when grown from stakes, or even faster when established from seed (Chadhokar, 1982). They are deciduous, glabrous and characterized by a spreading or pyrimidal crown of foliage borne on long, irregular, feathery branches that often curve downwards. They seasonally bear numerous large, showy, pink to light purplish, pea-shaped flowers which often appear before the leaves at the end of the dry season. The pods are glabrous, blackish at maturity and measure up to 14 cm long and 1.5 cm wide (Smith and van Houtert, 1987).
Native to tropical America and the West Indies, Gliricidin species now enjoy worldwide distribution as multipurpose trees suitable for use as living fences, for shade and soil improvement, as fuelwood and pole material, and as sources of high-protein fodder and browse. Gliricidia establishes well in the tropics: it is found in lowlands and at altitudes of up to 2000 m. Although it is best suited to the wetter areas in the humid tropics where it is widely cultivated, it is native to the drier parts of Central America. Its considerable drought tolerance is due to a policy of avoidance, since it drops its leaves and becomes dormant during the driest parts of the year. The plant thrives on relatively acid, infertile soils and shows some tolerance of short-term waterlogging, conditions which are unsuitable for some of the other popular tree legumes such as Leucaena leucocephala (Chadhokar, 1982).
Most published data on Gliricidia refer to G. septum, or to G. maculata. There is some confusion in the literature regarding the taxonomy, since authors such as Allen and Allen (1981) consider these taxa to be synonymous, while Wiersum and Dirdjosoemarto (1987) regard them as separate species. NFTA (1989) describes G. maculata as being native to the Yucatan Peninsula and having white flowers, rounded leaflets and relatively small pods and seeds. In contrast, G. septum is considered to be native to the Pacific coast of Mexico and Central America and has pale coloured flowers, more pointed leaves and larger seeds and pods. G. ehrenbergii comes from highland areas with altitudes of 1500-2000 m. It is a shrub growing to 3 m in height, with deep red-to-purple flowers and small rounded leaflets. This review will concentrate on G. septum and G. maculata, although most published reports probably refer to the same species, due to taxonomic confusion.
Because of the ease with which the species may be vegetatively propagated, it is likely that most of the work carried out in a number of distinct countries has concentrated on a narrow gene base.
Fodder characteristics
The foliage of Gliricidia has been more or less accepted throughout the tropics as a source of high quality fodder. This is despite some traditional beliefs that the plant is toxic at the flowering stage, particularly to non-ruminants (Carew, 1983). Seeds or powdered bark are mixed with rice for use as a poison for rats and mice (Uphof, 1968), while the leaves are reputedly toxic to dogs and horses but not to cows and goats (Blohm, 1962). Some non ruminants, however, are able to tolerate it, at least at low levels of intake, since it has been successfully used in Venezuela as a replacement for Medicago sativa (lucerne, alfalfa), as a yolk colourant in maize based diets for laying hens (Ford, 1987). Levels of 2-4% of Gliricidia leaf meal are recommended for poultry diets (NFTA, 1989). As a supplement for fish, Gliricidia meal has compared favourably with other leguminous fodders (Raj and Kutty, 1984).

Table 1 - Proximate and fibre
analyses of leaves and bark of Gliricidia sepium
Reports vary on the toxicity of G. septum. The flowers and leaves are eaten by humans where it grows but they may be detoxified by the cooking process (Martin and Ruberte, 1975). It was claimed by Neal (1948) that plants grown in Hawaii were not poisonous. Despite the conflicting reports, it has been successfully used for feeding ruminant livestock in many tropical countries of the world.
As a browse plant, the shoots and sprouts, especially the tender twigs, together with the fruits, pods and stems, are cropped by domestic and wild animals. Fodder is also cut at varying intervals and carried to provide a high protein supplement to small and large ruminants. Its use in alley cropping is widely reported (ILCA, 1988). The trees can withstand grazing and lopping and can be trimmed to a height of 1-1.5 m to serve as living fence posts and to provide limited forage within reach of browsing animals.
Data concerning browse productivity are few and variable and must be viewed with caution, since the age, species and size of the trees as well as the prevailing agro-climatic conditions can have considerable influences on foliage yields. ILCA (1988) cited a yield of 4475 kg/ha leaf dry matter (DM) over a 19-month period, equivalent to an annual DM yield of around 2800 kg/ha for the humid region of Nigeria,. In a similar area, a threeyear old crop of Gliricidia yielded 1131-2703 kg/ha DM over a five month period, equivalent to annual DM production of 2700 to 6500 kg/ha. Management practices will also have an effect on yield.
Data on the variation in yield with season are even scarcer and presumably more variable than those for annual production. Oakes and Skov (1962) reported monthly yields of 990 kg/ha for Gliricidia in the dry season, compared with 1480 kg/ha in the wet season. During times of stress, Gliricidin will shed its leaves, a characteristic which detracts from its usefulness as a standing reserve of high quality fodder to be carried over for feeding as a dry season supplement for lower quality roughages.
The yield of fresh forage of the whole tree is closely related to the fresh weight of a single branch in both Gliricidia and Leucacna. This gives a rapid, largely non-destructive method for estimating forage yields of intensively managed plantations (Sumberg, 1984).
Typical data for proximate and fibre analyses of G. septum are shown in Table 1. In general, they indicate good feeding value. NFTA (1989) confirmed this conclusion, suggesting that the foliage contained high levels of crude protein and calcium but low levels of phosphorus. Amino acid profiles (Chadhokar, 1982) compared favourably with herbaceous legumes such as Medicago saliva.
In a review, Smith and van Houert (1987) concluded that, in common with many legumes, G. septum contained sufficiently high levels of most minerals (except phosphorus and copper) to meet tropical requirements and it would make an excellent feed during the dry season, when protein and mineral deficiencies commonly occur. These comments apply equally to fresh foliage or to material conserved as dried leaf meal. Numerous reports consider the benefits of Gliricidia in terms of animal production, often as a supplement to poor quality diets (Ash, 1989). The foliage is highly degradable in the rumen (Minor and Hovell, 1979) and it is well digested (Wiersum and Dirdjosoemarto, 1987). Although rarely offered as a sole feed, Gliricidia should serve to increase the digestibility of the diet when used to supplement low quality feeds (NFTA, 1989).
Despite numerous reports to the contrary (e.g. NFTA, 1989), Smith and van Houtert (1987), reviewed performance trials with both small and large ruminants, and concluded that while the feeding of Gliricidia had no adverse effect on growth and reproductive performance, supplementation did not improve intake of the basal diet. When readily consumed, the bulky Gliricidia quickly distended the rumen, cutting down on the intake of the basal ration. In contrast to this, Ash (1989) considered that feeding small quantities of Gliricidia leaf to goats would significantly increase total DM intake when compared with mature tropical grass offered as a sole feed. Similarly, Chadhokar and Katharaju (1980) and Chadhokar and Lecumwasam (1982) reported increased levels of total intake when Gliricidia supplemented diets for dairy cattle and sheep respectively. These contrasting observations may be due, at least in part, to differences between species, or even between provenances, since little work has been done to characterize and stabilize varieties of Gliricidia.
Crushed Gliricidia leaves have been associated with a strong odour and this has resulted in palatability being described as only satisfactory to sheep (Nochebuena and O'Donovan, 1986) and unpalatable to rabbits (Raharjo and Cheeke, 1985). However, Chadhokar and Lecamwasam (1982) described Gliricidia as both acceptable and palatable to dairy cattle. While differences between provenances may explain some of the variability in reported animal acceptance, wilting for 24 hours between cutting and feeding appears to improve intake by reducing the odour of the foliage. The same effect can be achieved by oven drying. It has been reported that animals which initially refused Gliricidia became accustomed to it in a few days, after which it was readily consumed. The presence of animals used to eating the foliage accelerated acceptance by others which were being introduced to it (Atta-Krah and Sumberg, 1987).
The rather conflicting data suggest that Gliricidia can be a useful supplement to low quality roughage diets, even if the improvement in animal performance is not great.
Anti-nutritive factors
Early reports concerning the role of Gliricidia as a forage refer to its toxicity and certainly the derivation of the generic name (glis = doormouse, caedo = kill) implies a toxicity to rodents. Furthermore, the powdered bark and seeds have been used as a rodent poison in the tropics and Skerman et al. (1988) confirmed that the roots were toxic to rodents and suggested that the leaves could poison horses.
Potentially toxic substances have been isolated from Gliricidia, including coumarins (Griffiths, 1962), hydrocyanic acid (Manidool, 1985), alkaloids (Glander, 1977), and tannins (Devendra, 1983).
Furthermore, it is suspected that Gliricidia may be a nitrate accumulator responsible for a 'cattle fall syndrome' in Colombia (Tiheebilcock, 1978).
However, whilst toxic effects have been well documented in non-ruminants, conclusive evidence in ruminants is lacking. Recent work (Ash, 1989) reported moderate condensed tannin levels in Gliricidia (20.5 g/kg DM), and yet the plant material was more degraded in the rumen than that of other plants of a similar tannin content. Condensed tannins may differ in type and behaviour between plant species. It also appeared that tannins could be altered by feed treatment. Oven-drying, a process by which animal acceptance has been increased, resulted in an apparent loss of tannins (Akin et al., 1989). Palatability could be improved by this process, although it could also have been modified by the destructive effect of heat on other plant components such as coumarins.
In conclusion, under practical feeding conditions, Gliricidin does not seem to be toxic to ruminants, although palatability may be a problem in some areas. It should be used with caution as a dietary supplement of diets for non-ruminants.
Management
Gliricidia tends to shed its leaves during the dry season and in Central America flowering and fruiting often take place while the trees are bare (February to April). In parts of Asia and the South Pacific, flowering is in the wet season and seed production is then often poor. Mature pods are strongly dehiscent, throwing their contents up to 25 m from the parent tree and creating problems for seed collection. The seeds show no dormancy, germinating readily as soon as moisture becomes available. If the opening rains are followed by a dry spell, many seedlings growing from naturally dispersed seed will die before they can become established. While there is considerable variation between individual trees, Gliricidia is not generally a heavy seeder (Salazar, 1986; Atta-Krah, 1987), although the seed maintains viability for well over a year when stored at 17°C and 50% humidity (NFTA, 1986). Trees grown from seed appear to produce stronger and deeper root systems than those which are vegetatively propagated, so are more suitable for use on sloping land, or where drought and high winds are potential problems.
Propagation by long stakes is easy, giving rise to the Caribbean common name of quickstick. Mature, leafless cuttings (6-12 months growth), 0.5-1.5 m long, produced a strike rate of some 85% when bark stripping was combined with regular watering (Grover, 1986). Chadhokar (1982) reported best results from fresh stakes 1.0-1.5 m in length and 3-5 cm in diameter, when about 15 cm of the stake was embedded in the soil. Coating the exposed ends with vaseline, wax, mud or polythene aided survival by reducing moisture loss.
The ease of vegetative propagation and the widespread use of Gliricidia as living fence posts established from long stakes, coupled with seed production difficulties, has probably resulted in the exploitation of only a small proportion of the existing germplasm. There are collections of seed held at international centres such as the Nitrogen Fixing Tree Association in Hawaii and the Oxford Forestry Institute in the UK (NFTA, 1989). Testing these seed resources should be given priority, although there would appear to be scope for further collection and selection from native populations in order to obtain provenances with desirable characteristics before the gene base becomes further eroded. Wider use of seed would enable superior individuals to be selected from both natural and artificially established populations.
In alley cropping, in order to leave adequate space for annual crops, it is common to plant the trees in rows 4-10 m apart on flat land, reducing the spacing to 2-3 m as the slope of the land increases. In either case, spacing within the row is 10-50 cm. The trees are pruned at the end of the first year and subsequently managed with periodic cutting of regrowth to prevent excessive shading of the interplanted crops. It has been reported that delaying the age at first cutting encourages root development. Chadhokar (1982) recommended one or two harvests per year for the first two or three years, followed by an 8-12 week cutting cycle (4-5 cuts/year) to maximize the yield of foliage. Two cuts during the wet season delayed leaf fall and flowering during the subsequent dry season (Simmonds, 1951). Reduction of the leaf canopy at this time also protected trees from the ravages of storms and hurricanes in the Caribbean.
Little work has been reported on tree spacings for forage production. Unlike Leucaena, which can be easily maintained as a hedge for direct grazing, Gliricidia does not respond well to repeated cutting close to ground level, although NFTA (1989) reported that it would tolerate a cutting height of 40-90 cm. Partly as a result of its use as living fence posts, it is common to see the trees growing with a single trunk to a height of at least 1.5 m, branching profusely above that height. This implies management under what would be basically a cutting regime and the optimum pattern and density in plantations would then be determined by the methods of harvesting and transporting the cut material. Densities in excess of 3000 trees/ha are probably rare in practice, although NFTA (1989) noted spacings of 1.0 x 0.25 m in protein banks and Wiersum and Dirdjosoemarto (1987) quoted spacings ranging from 1 x 1 m to 2.5 x 2.5 m for woodlots in Indonesia. In fence lines, spacings of 0.5-3.0 m are common.
Under a cutting regime, some of the cut material is usually fed fresh to livestock, although sun-dried leaf material is easy to store for later use. Dried foliage is well accepted. Gliricidia lacks the fermentable carbohydrates necessary to make high quality silage unless it is ensiled with molasses and formic acid additives (Kass and Rodriguez, 1987). Wilting also improves the quality of the silage.
At wide spacings of 10 x 10 m in a parkland grazing system, Gliricidia can provide shade and foliage to animals and small quantities of nitrogen to the companion grass species without reducing grass yield through excessive shading.
Alternative uses
Gliricidia is a truly multipurpose tree with a variety of uses complementing its role as a source of fodder. Its ease of establishment from large vegetative cuttings makes it suitable for living fences where it can also act as a windbreak, or as a support for climbing crops such as yam, pepper, vanilla and passion fruit. The excessive new tree growth is browsed or lopped for fodder, while the blossoms provide pollen and nectar for honeybees during the short flowering period.
Gliricidia serves a variety of soil protection and improvement purposes. As well as the fencing and support roles noted above, it is often interplanted with plantation crops as shade and to provide green manure or mulch. It has long been used as a shade tree in tropical plantations of tea, coffee and cocoa, with benefits including ease of husbandry and propagation, good coppicing, a canopy structure that permits a desirable level of shade and a deep, noncompetitive root structure (Budowski et al., 1984). Associations with annual crops provide both shade and nutrients. It is this further role as a source of nutrients in alley cropping that makes it ideally suited to the improvement of farming systems through both maintenance of soil fertility for crop production and provision of high protein feed for ruminants.
Alley farming systems, developed by the International Livestock Centre for Africa (ILCA) and the International Institute of Tropical Agriculture (IITA) for the humid regions of the developing world, employ foliage to both improve the soil and to provide animal feed. They have led to increased maize yields, due to the benefits of nitrogen fixation by the root nodules and by the addition of nutrients supplied as either green manure or mulch. Timely pruning is essential since failure to remove superfluous foliage can cause decreased crop yields due to excessive shading, together with increased risk of trees being blown over during heavy storms. Foliage incorporated into the soil as green manure, rather than applied to the surface as mulch, results in faster leaf decomposition and nutrient release. In drier areas and areas subject to soil erosion, mulching may be more appropriate to reduce soil temperature and increase soil moisture retention. Wilson et al., (1986) suggested that shrubs may be more suitable than herbaceous legumes for soil restoration in the humid tropics. The nitrogen released from decomposing plant tissue may be more important than that exuded from roots in such environments.
In numerous agroforestry systems, nutrient release by leaf decomposition is regarded as the most important factor in the restoration of soil fertility. Roskoski et al., (1982) measured rhizobial activity in situ in a 20-year-old stand of G. septum and estimated annual nitrogen fixation to be only 13 kg/ha/year. In comparison, Yamoah et al., (1985) reported that prunings released 252 kg nitrogen/ha/year when G. septum was intercropped with maize. This would represent nitrogen both from the soil and from rhizobial fixation. While Gliricidia is known to form nodules, there is little information available regarding the efficiency of nitrogen fixation under normal farm conditions.
As a source of nitrogen and organic matter, Gliricidia is often added to rice fields at mudding up. Gliricidia compost was compared with cow, pig, goat, and poultry manure for effects on rice growth. The highest grain yield, 5.7 t/ha, resulted from the application of both Gliricidia and pig manure (Joseph and Kurakose, 1985).
A balance should be sought between the use of the foliage as mulch, as green manure or as fodder, depending upon the nature of the farming system. The deep rooting habit of leguminous trees is an important feature in the prevention of soil erosion, especially with increased cultivation of more precarious hillsides.
Aken'Ova and Atta-Krah (1986) noted additional benefits of improved weed control in alley cropping, while Obando (1987) indicated a possible allelopathic effect of Gliricidia on the common weeds Bidens pilosa and Melanpodium perfoliatum in Costa Rica. Sclerotial viability of Rhizoctonia solani, a pathogen of rice, is reduced by the addition of green Gliricidia leaves to the soil (Lakshmanan and Nair, 1984). Hot water infusions of the leaves are used as pesticides against external parasites of livestock, including dogs, and chopped leaves placed in the nests serve a similar purpose in poultry husbandry (NFTA, 1989).
Hot compresses of Gliricidia leaves, bark and roots are used to treat wounds, warts and bites, and skin rashes are cured by bathing in hot water infusions of the leaves (NFTA, 1989).
The wood of Gliricidia is hard and heavy (specific gravity 0.75) with a coarse texture and irregular grain. It is not easy to work but finishes smoothly and is extremely durable. It is used to fashion agricultural implements and small carpentry items which show great resistance to termites and decay. While Soetrisno et m., (1984) produced satisfactory wood pulps in Indonesia, Yantasuth et al., (1985) considered G. sepium unsuitable for pulping in Thailand.
A calorific value of 4900 kcal/kg is commonly cited for Gliricidia when used as a fuelwood, and it has been described as an outstanding fuelwood tree by Brewbaker et m., (1982)
References and further reading
Two important references which report the level of current knowledge on Gliricidia septum have been recently published by the Nitrogen Fixing Tree Association in Hawaii. Where appropriate, individual contributions to these volumes have been noted in the alphabetical listing of references below. These publications are:
NFTA (1987) Gliricidia septum (Jacq.) Walp.: Management and Improvement. Proceedings, Workshop held at CATIE, Turrialba, Costa Rica, rune, 1987. Special Pub. 87-01. Hawaii, USA: Nitrogen Fixing Tree Association.
NFTA (1989) Gliricidia Production and Use. GLOVER, N. (ed). Hawaii, USA: Nitrogen Fixing Tree Association.
Alphabetical listing
AHN, J.H., ROBERTSON, B.M., ELLIOT, R., GUTI ERIDGE, R.C. and FORD, C.W. (1989) Quality assessment of tropical browse legumes: tannin content and protein degradation. Animal Feed Science and Technology, 27: 147-156.
AKEN'OVA, M.E. and ATTA-KRAH, A.N. (1986) Control of spear grass (Imperata cylindrica (L.) Beauv.) in an alley cropping fallow. Nitrogen Fixing Tree Research Reports, 4: 27-28.
ALLEN, O.N. and ALLEN, E.K. (1981) The Leguminosue: A Source Book of Characteristics, Uses and Nodulation. Madison, USA: The University of Wisconsin Press.
ASH, A.J. (1989) The effect of supplementation with leaves from the leguminous trees Sesbania grandiflora, Albizia chinensis and Gliricidia septum on the intake and digestibility of guinea grass hay by goats. Animal Feed Science and Technology, 28: 225-232.
ATTA-KRAH, A.N. (1987) Flowering and seed production of Gliricidia septum. In: Gliricidia septum (Jacq.) Walp.: Management and Improvement. Proceedings, Workshop held at CATIE, Turrialba, Costa Rica, June, 1987. Special Pub. 87-01. pp 20-28. Hawaii, USA: Nitrogen Fixing Tree Association.
ATTA-KRAH, A.N. and SUMBERG, J.E. (1987) Studies with Gliricidin septum for crop-livestock production systems in West Africa. In: Gliricidia septum (Jacq.) Walp.: Management and Improvement. Proceedings, Workshop held at CATIE, Turrialba, Costa Rica, June, 1987. Special Pub. 87-01. pp 31-43. Hawaii, USA: Nitrogen Fixing Tree Association.
BLOHM, H. (1962) Poisonous Plants of Venezuela. Cambridge, USA: Harvard University Press.
BREWBAKER, J.L. (1986) Leguminous trees and shrubs for Southeast Asia and the South Pacific. In: Forages in Southwest Asian and South Pacific Agriculture BLAIR, G.J., IVORY, D.A. and EVANS, T.R.(eds). ACIAR Proceedings No 12, Canberra, Australia. pp 43-50.
BREWBAKER, J.L., MACDICKENS K., and VAN DEN BELDT, R., (1982) Tropical nitrogen-fixing fuelwood trees. In: Energy Conservation and Use of Renewable Energies in the Bio-industries. VOGT, F.(ed). pp 108-119 Oxford, UK: Pergamon Press.
BUDOWSKI, G., KASS, D.C.L. and RUSSO, R.O. (1984) Leguminous trees for shade. Pesquisa Agropecuaria Brasileira, 19: 205-222.
CAREW, B.A.R. (1983) Gliricidia septum as a sole feed for small ruminants. Tropical Grasslands, 17(4): 181-184.
CHADHOKAR, P.A. (1982) Gliricidia maculata - A promising legume fodder plant. World Animal Review, 44: 36-43.
CHADHOKAR, P.A. and KANTHARAJU, H.R. (1980) Effect of Gliricidia maculata on growth and breeding of Bannur ewes. Tropical Grasslands, 14: 78-82.
CHADHOKAR, P.A. and LECAMWASAM, A. (1982) Effect of feeding Gl�ricidia maculata to milking cows: a preliminary report. Tropical Grasslands, 16: 46-48.
DEVENDRA, C. (1983) Physical treatment of rice straw for goats and sheep and the response to substitution with variable levels of cassava (Manihot esculenta Crantz.), leucaena (Leucacna leucocephala) and gliricidia (Gliricidia maculata) forages. MARDI Research Bulletin, 11(3): 272-290.
FALVEY, J.L. (1982) Gliricidia maculata - a review. International Tree Crops Journal, 2(1): 1-14.
FORD, L.B. (1987) Experiences with Gliricidia septum in the Caribbean. In: Gliricidia septum (Jacq.) Walp.: Management and Improvement. Proceedings, Workshop held at CATIE, Turrialba, Costa Rica, June, 1987. Special Pub. 87-01. pp3-7. Hawaii, USA: Nitrogen Fixing Tree Association.
GLOVER, N. (1986) Vegetative propagation of Gliricidia septum. Nitrogen Fixing Tree Research Reports, 4: 62-63.
GLANDER, K.E. (1977) Secondary compounds and the feeding behaviour of leaf-eating primates. American Journal of Physical Anthropology, 47: 133.
GRIFFITHS, L.A. (1962) On the co-occurrence of coumarin, O-coumaric acid and melilotic acid in Gl�ricidia septum and Dipteryx odorata. Experimental Botany, 13: 169-175.
ILCA (1988) Annual Report. Addis Ababa, Ethiopia: International Livestock Centre for Africa.
JOSEPH, P.A. and KURIAKOSE, T.F. (1985) An integrated nutrient supply system for higher rice production. International Rice Research Newsletter, 10(2): 22.
KASS, M.L. and RODRIGUEZ, G. (1987) Preliminary studies on silage making from Gliricidia septum (Madero Negro). In: Gliricidia septum (Jacq.) Walp.: Management and improvement. Proceedings, Workshop held at CATIE, Turrialba, Costa Rica, rune, 1987. Special Pub. 87-01. pp201-204. Hawaii, USA: Nitrogen Fixing Tree Association.
LAKSHMANAN, P. and NAIR, M.C. (1984) Effect of soil amendments on the viability of sclerotia of Rhizoctonia solani in soil. Madras Agricultural Journal, 71(8): 526-529.
MANIDOOL, C. (1985) Utilisation of tree legumes with crop residues as animal feeds in Thailand. In: Relevance of Crop Residues as Animal Feeds in Developing Countries. Proceedings, International Workshop held at Khoen Kaen, Thailand. pp 249-269.
MARTIN, F.W. and RUBERTE, R.M. (1975) Edible Leaves of the Tropics. Mayaguez, Puerto Rico: Antillean College Press.
MINOR, S. and HOVELL, D.F.D. (1979) Rate of rumen digestion of different protein sources using the in vivo nylon bag technique with cattle fed sugar cane. Tropical Animal Production, 4: 105-106.
NEAL, M.C. (1948) In Gardens of Hazouii. Special Publication No.40. Honolulu, USA: Bernice P. Bishop Museum.
NFTA (1986) Gliricidia: its name tells its story. NFT Highlights 86-06. Hawaii, USA: Nitrogen Fixing Tree Association.
NOCHEBUENA, G. and O'DONOVAN, P.B. (1986) The nutritional value of high-protein forage from Gliricidia septum. World Animal Review, 57: 48-49.
OAKES, A. and SKOV, O. (1962) Some woody legumes as forage crops for the dry tropics. Tropical Agriculture 39(4): 281-287.
OBANDO, L. (1987) Potencial alelopatico de Gliricidia septum (Jacq.) Walp. sobre los cultivos de maiz y frijol y las malezas predominantes (abstract). In: Gliricidia septum (Jacq.) Walp.: Management and Improvement. Special Pub. 87-01. pp 59-60. Hawaii, USA: Nitrogen Fixing Tree Association.
PATIL, S.D. and PRASUNAMMA, K. (1986) A preliminary evaluation of nodules from six nitrogen fixing trees. Nitrogen Fixing Tree Research Reports, 4: 20-21.
RAHAJO, Y.C. and CHEEKE, P.R. (1985) Palatability of tropical tree legume forage for rabbits. Nitrogen Fixing Tree Research Reports, 3: 31-32.
RAJ, S.P. and KUTTY, M.N. (1984) Food conversion efficiency and nitrogen balance in Cirrhinus mrigala fingerlings fed on three pelleted feeds compounded with wild legumes. Journal of the Indian Institute of Science, 65(�): 59-64.
ROSKOSKI, J.P., MONTANO, J., VAN KESSEL, C. and CASTILLEJA, G. (1982) Nitrogen fixation by tropical woody legumes: potential source of soil enrichment. In: Biological nitrogen fixation technology for tropical agriculture. GRAHAM, P.H. and HARRIS, S.C.(eds). pp 447-454. Cali, Colombia: Centro Internacional de Agricultura Tropical (CIAT).
SALAZAR, R. (1986) Genetic variation in seeds and seedlings of 10 provenances of Gliricidia septum (Jacq.) Steud. Forest Ecology and Management, 16: 391-401.
SIMMONDS, N.W. (1951) Notes on field management at the Botany Department of the Imperial College of Tropical Agriculture. Tropical Agriculture (Trinidad), 28(1-6): 70-75.
SKERMAN, P.J., CAMERON, D.G. and RIVEROS, F. (1988) Tropical Forage Legumes. 2nd. edn. Rome: Food and Agriculture Organization.
SMITH, O.B. and VAN HOUTERT M.F.J. (1987) The feeding value of Gliricidia sepium: a review. World Animal Review, 57: 57-68.
SOETRISNO, SAYUTI, MULIAH and EDDI, M. (1984) Pulping of Gliricidia maculata. Berita Selulosa 20(1): 3-12.
SUMBERG, J. (1984) Alley farming with Gliricidia septum. Germplasm Evaluation and Planting Density. Ibaban, Nigeria: ILCA Small Ruminant Program.
TIHEEBILCOCK, P.E. (1978) Nitratos totales en plantas y agues
estancadas de Cordoba y Sucre como factores causantes de "la caida del ganado".
Revista
del Instituto Colombiano de Agricultura (Bogota), 13: 567-574.
UPHOF, J.C.T. (1968) Dictionary of Economic Plants. 2nd edn. Lehre, W. Germany: V.J. Von Cramer.
WIERSUM, F. and DIRDJOSOEMARTO, S. (1987) Past and current research with Gliricidia in Asia. In: Gliricidia sepium (Jacq.) Walp.: Management and Improvement. Proceedings, Workshop held at CATIE, Turrialba, Costa Rica, rune, 1987. Special Pub. 87-01. pp 20-28. Hawaii, USA: Nitrogen Fixing Tree Association.
WILSON, G.F., KANG, B.T. and MULONGOY, K. (1986) Alley cropping: trees as sources of greenmanure and mulch in the tropics. Biological Agriculture and Horticulture, 3-213): 251-267.
YAMOAH, C.F., MULONGOY, K. and AGBOOLA, A.A. (1985) Decomposition and nitrogen contribution by prunings of selected legumes in alley cropping systems. In: Biological Nitrogen Fixation in Africa. SSALI, H. and KEYA, S.O.J. (eds.) pp482-485. Nairobi, Kenya: MIRCEN, University of Nairobi.
YANTASATH, K., SUPATANAKUL, W., UNGVICHIAN, I., CHAMSAWAD, S., CHANTRASIRI, S., PATANAVIBUL, S., HYAKITKOSOL, C., PROMPETCHARA, S., PITHAKARNOP, N. and CHALERMKLIN, P. (1985) V. Pulping and paper making characteristics of fast growing trees. Nitrogen Fixing Tree Research Reports 3: 54-56.
Other titles in the Use of Trees by Livestock series include:
1 Prosopis
2 Acacia
4 Anti-nutritive factors
5
Quercus
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Use of Trees by Livestock (NRI, 1994, 160 p.)
Use of Trees by Livestock: Anti-Nutritive Factors
R.T. Paterson
Acknowledgements
© Crown copyright 1993
The Natural Resources Institute (NRI) is an internationally recognized centre of expertise on the natural resources sector in developing countries. It forms an integral part of the British Government's overseas aid programme. Its principal aim is to alleviate poverty and hardship in developing countries by increasing the productivity of their renewable natural resources. NRI's main fields of expertise are resource assessment and farming systems, integrated pest management, food science and crop utilization.
NRI carries out research and surveys; develops pilot-scale plant, machinery and processes; identifies, prepares, manages and executes projects; provides advice and training; and publishes scientific and development material.
Short extracts of material from this publication may be reproduced in any non-advertising, nonprofit-making context provided that the source is acknowledged as follows:
Paterson, R.T. (1993) Use of Trees by Livestock 4: Anti-nutritive Factors Chatham, UK: Natural Resources Institute.
Permission for commercial reproduction should be sought from:
The Head, Publishing and Publicity Services, Natural Resources Institute, Central Avenue, Chatham Maritime, Kent, ME4 4TB, United Kingdom.
No charge is made for single copies of this publication sent to the governmental and educational establishments, research institutions and non-profitmaking organizations working in countries eligible for British Government Aid. Free copies cannot normally be addressed to individuals by name but only under their official titles.
Natural Resources Institute ISBN: 0 85954 335-8 Price £2.00 LG4
This booklet is printed on, paper produced using the residues from sugar-cane processing.
Foreword
The importance of trees and shrubs in the feeding of animals in the tropics and sub-tropics has long been recognized by livestock owners. In arid areas where the growth of herbaceous plants is limited by lack of moisture, leaves and edible twigs of trees and shrubs can constitute well over 50% of the biomass production of rangeland. At high altitudes, tree foliage may provide over 50% of the feed available to ruminants in the dry season, branches being harvested and carried to the animals. Even in regions of higher rainfall where grass supplies the major proportion of the dry matter eaten by ruminants, tree leaves and fruits can form an important constituent of the diet, particularly for small ruminants.
In the last two decades interest in the planting of trees as a source of feed for livestock has been encouraged by workers in research and development, but in contrast to the hundreds of indigenous species which are used as fodder, attention has focussed on a limited number of introduced species. Thus there are many publications reporting the chemical composition of Leucaena leucocephala leaves and suggesting management strategies for utilization of the tree for fodder, but it is more difficult to find information on alternative genera which might be equally, or more, appropriate.
The aim of this series of publications is to bring together published information on selected genera of trees which have the potential to increase the supply of fodder for ruminants. Each booklet summarizes published information on the fodder characteristics and nutritive value of one genus, with recommendations on management strategies, where available. Since the leaves of woody species frequently contain secondary compounds which may have an anti-nutritional or toxic effect, this booklet summarizes the effects of a number of these compounds. It is hoped that the booklets will provide useful resource material for students, research and extension workers, interested in promoting the use of trees as a source of fodder for ruminants.
Further copies of this booklet or others in the series can be obtained by writing to Publishing and Publicity Services at the Natural Resources Institute.
Margaret Gill Livestock Production Programme
Summary
Woody plants contain a wide array of anti-nutritive factors which can have either repellent or adverse effects on livestock. While physical features of plants will serve only to limit intake and hence production, chemical components may lead to stock losses through poisoning. Manifestations of toxicity will depend on the level of toxin in the plant (which itself varies with the plant part, the variety, the species, the season of the year and the conditions under which it has grown), the level of intake of the feed and the extent and rate of metabolism of the toxic constituents within the animal. The simultaneous occurrence of several toxic substances in the same feed creates difficulties in establishing a hierarchical order of potency and complicates attempts to detoxify the feed by simple, cost-effective means.
There is a need for further research to elucidate some of the problems involved in the utilization of foliage from trees and shrubs. At present, perhaps the greatest need is for the development of laboratory tests which accurately reflect animal performance in the presence of anti-nutritional factors such as tannins and alkaloids. This will require extensive collaboration between chemists, agronomists and animal nutritionists in order to establish correlations under a wide range of conditions of both plant and animal growth.
Introduction
Anti-nutritive factors are those physical and chemical features of plants potentially available for grazing or browsing, which result in lower levels of animal productivity than would be expected from proximate and mineral analyses of the foliage. In general, they serve to protect the plants from excessive damage by being feeding deterrents to herbivores. Many factors appear to be the natural result of co-evolution of the plants with herbivorous animals (Levin and York, 1978; Brewbaker, 1989). Others have probably evolved as protection mechanisms against insect pests and plant diseases (D'Mello, 1982), in which case their effects on higher animals may be coincidental.
Many features of plants are capable of reducing their acceptability to animals and they range from the purely physical, to symbiotic relations and chemical reactions. Plants can be rendered unattractive as forage by virtue of their effects on any of the senses of the animal. Acacia Iysiphloia has sticky foliage, and A. coriacea gives off an offensive odour, both of which are repellent to livestock and result in reduced acceptance of the feed (Fox, 1987). Alternatively, physiological effects on digestion and metabolism can also reduce voluntary intake by the grazing animal, with a consequent loss of productivity. Many of the chemical effects are caused by potentially toxic substances. Under normal grazing conditions intake may be insufficient to result in stock losses, but animal productivity is often reduced even in the absence of physical symptoms of toxicity. Deaths usually occur in such cases only when animals are hungry and access to alternative fodder is limited.
The study of anti-nutritive factors is complicated by the frequent occurrence of more than one toxic compound in a single feed source. This creates difficulty in the establishment of a hierarchical order of potency. The toxic principles may also act synergistically. The levels of deleterious substances vary with plant part, species, cultivar, growing conditions and season of the year and post-harvest treatments such as drying, soaking, autoclaving and germination of seed material. Extrapolation from case to case must therefore be made only with great care.
Shrubs and trees exist which are highly toxic at all times of the year and under almost all circumstances. They contain phytotoxins, sometimes known as toxalbumins (Kingsbury, 1964). These complex molecules, similar in both structure and physiological reaction to bacterial toxins, can kill at extremely low levels of intake, as with Robinia pseudoacacia (black locust) and Rinicus communis (castor bean). These plants are generally avoided by livestock, except when severe nutritional stress makes feed selection difficult, or when toxic plants are cut and mixed with other, acceptable fodder. Animals which have never been exposed to the poisonous plants are at greater risk than those which are accustomed to them.
Such plants and their toxic principles are beyond the scope of this review, which deals only with those anti-nutritive factors commonly found in shrubs and trees with potential for feeding domestic livestock.
Acceptability of plants
Physical features Physical features of plants generally affect their acceptability to livestock and consequently the level of voluntary feed intake. Several aspects of the acceptability of fodder are purely physical in nature and are considered below.
Growth form
This is the most obvious physical deterrent to consumption by herbivores. Many herbaceous species avoid being grazed through a creeping growth habit. While animals such as horses and sheep can graze close to ground level, cattle are limited in their ability to defoliate prostrate plants such as Desmodium triflorum. Most trees will rapidly grow out of the reach of domestic grazing animals unless they are suitably managed. Species such as Leucaena leucocephala will readily regrow after severe cutting or defoliation, even to ground level, while others such as Sesbania spp. are much less tolerant of complete defoliation (Skerman et al., 1988).
Thorns and spines
Within any given genus there may be both armed and unarmed species, but Acac�a, Caesalpinin, Erythrina, Mimosa and Prosopis are all recognized for their dangerous thorns and spines (Allen and Allen, 1981). The presence of these features and their distribution on the plant are of great importance in determining their usefulness as a browse species for different classes of livestock.
Selective grazers such as goats and camels can consume foliage that is unacceptable to cattle and sheep; and while sheep make use mainly of fallen leaves and pods of Prosopis tamarugo in Chile, goats are, to some extent, able to avoid the spines while browsing the green foliage (Habit et al., 1981). In Australia, Acacia polyacantha has spines that protect it from most grazing animals (Skerman et al., 1988), while in Africa, species such as A. detinens, A. karoo and A. tortilis form impenetrable thickets which can impale game (Allen and Allen, 1981).
Myrmecophily is an obligatory, symbiotic association between ants and plants, the best known example being the habitation of swollen, stipular Acacia thorns by ants of the genus Pseudomyrmex. The ants feed on the enlarged foliar nectaries and modified leaflet tips of the plant. In return, they protect it from defoliation by insects and herbivorous animals by swarming to attack when the tree is disturbed. Approximately nine neotropical species of Acacia share non-specific, obligatory relationships with some five ant species (Allen and Allen, 1981). The ants, amongst the most ferocious in the world, present a formidable defensive barrier to the grazing animal.
Other
Cattle generally show a marked preference for grass and soft herbage rather than tough, sclerophyllous leaves. This characteristic is shared, to some extent by sheep but probably not by goats. Such a preference, however, often assumes importance only when animals are presented with a choice. In arid and semiarid regions, or at particularly dry times of the year, the choice is often between consuming unattractive leaf material or going hungry. Foran (1984) noted that in central Australia, Acacia aneura and A. kempeana contributed up to 40% of the diet for cattle during the hardest times of the year, but when grass was abundant, the level normally fell to less than 20%. Thus, while the shrubs are clearly not the favourite cattle forage in this region, they are consumed in large quantities when the preferred species are not available.
Chemical features
The leaves of woody plants tend to contain more anti nutritive components (plant secondary compounds) than those of herbaceous plants. This is particularly true of phenolics (Bate-Smith, 1962), although there is a wide range of chemical components capable of having adverse effects on herbiverous animals.
Phenolic compounds
Plant materials contain a wide range of phenolics from low molecular weight, metabolizable phenols to the much larger polyphenols, a group which, with molecular weights in the approximate range of 5003000, includes the tannins. Phenolic compounds are commonly found in the leaves, bark, fruit and twigs of trees and are thought to confer resistance to both insect and fungal attack (D'Mello, 1982). Much has been written about the anti-nutritional effect of tannins on herbivores including recent reviews from McLeod (1974), Kumar and Singh (1984), Mangan (1988) and Kumar and Vaithiyanathan (1990).
Chemically, tannins are complex phenolic polymers containing aliphatic and phenolic hydroxyl groups and in some cases, carboxyl groups. The term tannin comes from the use of plant extracts in the tanning of leather which bind protein to form insoluble tannin-protein complexes. They are generally classified into two structural types, depending on their chemical properties.
Hydrolyzable tannins are mainly found in fruit pods and plant galls (Liener, 1980). They have a central carbohydrate core and can be degraded by chemicals or enzymes into a sugar residue and a phenol-carboxylic acid which can then be absorbed through the gut wall of the animal. These can be further subdivided depending on the nature of the phenol-carboxylic acid component into, for example, gallotannins and ellagitannins, which can influence feed intake by affecting the palatability of the feed. They may also have a toxic effect on animals (Mangan, 1988).
Condensed tannins (procyanidins or proanthocyanidins) have no carbohydrate core and are usually derived from condensation of flavonoid precursors. They cannot be degraded by enzymes and are therefore unlikely to pass through the gut wall. They are the principal tannins of forage legumes and they usually exist in the plant as leucoanthocyanins. The formation of protein-tannin complexes may depress the nutritive value of fodder by reducing both voluntary feed intake and digestibility. The acceptability of browse is clearly a complex matter but it appears to be related to the concentration of condensed tannins although there may be a threshold possibly at about 5%, below which there is no measurable effect (McNaughton, 1987).
Tannins mainly affect livestock by forming complexes with, and thus precipitating, proteins in the gut. Lower molecular weight polyphenolics in feeds, or products of tannin degradation, could also have an effect. Tannins provoke an astringent reaction in the mouth of the animal (Bate-Smith, 1973), probably by reacting with proteins in the saliva and the mucous epithelium, to impair lubrication in both the mouth and the oesophagus. They may inhibit digestibility by forming relatively indigestible protein-tannin complexes and by directly inhibiting digestive enzymes and micro-organisms. The effect on rumen microflora appears to be a result of the formation of complexes with proteins in the cell wall.
Tannins are assumed to reduce voluntary feed intake, either by their astringent effect, or by reducing protein digestibility and absorption but the exact mechanism is poorly understood and results are conflicting. Tannic acid added to the feed does not always produce the results ascribed to natural tannins. However, many factors interact to control voluntary intake (Forbes, 1986) and thus the response to individual factors will vary according to the specific circumstances.
There is a mechanism which protects animals from the potentially toxic effects of tannins and this is demonstrated by greater tolerance when either tannic acid or naturally occuring tannins are ingested orally rather than when administered by subcutaneous injection. At high levels of feed intake, tannins react with both muco-proteins and the outer cellular layer of the digestive tract. This appears to alter the permeability of the gut wall resulting in gastritis, intestinal irritation and constipation. Under suchconditions, tannins may be absorbed by the animal and result in liver and kidney damage if the physiological detoxication mechanisms are inadequate to cope with the influx.
The tolerance to tannins varies between animal species. The saliva of some species such as deer, rodents and goats, appears to contain proline-rich proteins, which may constitute a first line of defence against tannins. Cattle, sheep and chickens appear to have less, or possibly no capacity to secrete these proteins (D'Mello, 1992). Poultry and horses have been shown to develop symptoms of toxicity when fed tannic acid at levels of about 2% of diet (dry matter basis) under experimental conditions, while ruminants are able to handle higher levels of the acid without adverse effects.
Although deaths in sheep and cattle have been attributed to the high levels of tannins found in the foliage of trees such as Quercus spp., McLeod, (1974) concluded: 'there is no evidence that forage tannins have any detrimental effect upon the grazing ruminant, even though tannins have been shown to reduce protein digestibility.' A more recent review (Kumar and Singh, 1984), however, implicated tannins in: low milk yields; a reduction in available sulphur; in toxic, degenerative changes in the intestine, liver, spleen and kidney; and the appearance of mucus in urine and fatal constipation. These authors suggested that tannin-rich fodder for ruminants should be restricted or fed with caution. They also pointed out that the risk to unconfined animals was generally small, since given a choice they will seldom consume enough tannin to suffer harm.
The literature abounds with conflicting reports on the effects of tannin content on animal performance. There are several possible explanations for the apparent contradict�ons.
(a) The choice of assay. There are a number of analytical techniques which have been used to estimate tannin content and these have been critically reviewed by Hagerman and Butler (1989). They are based on measures of either chemical groups and structures (e.g. based on Folin, or Prussian blue reagents, vanillin or butanol) or biochemical activity (e.g. protein precipitation tests). Different assays can result in widely varying estimates of tannin content, since they are designed to measure different characteristics. Ahn et al., (1989) reported that leaf dry matter of Acac�a angustissima contained up to 6.5% tannin when assayed by the vanillin-HCI method, whereas no tannins could be detected by the butanolHCl technique. There was also poor correlation in some browse species between nitrogen digestion and total condensed tannin content as measured by either vanillin or butanol-HCl assays. Clearly determinations made using different techniques are not necessarily comparable. Hagerman and Butler (1981) noted evidence that the protein binding capacity of tannins could be highly specific to certain proteins.
(b) The presence of other factors. More than one anti-nutritive factor can be present in a single feed and this can confound results. Tanner et al., (1990) suggested that the poor performance of sheep fed on pods of Acacia sieberiana was due to the presence of both tannins and cyanogenic glycosides.
(c) Seasonal variation. Both the concentration and the type of phenolic compounds within the plant are subject to continual change. Van Hoven (1985) described the rapid mobilization of phytochemicals in response to defoliation by animals. As leaves and fruits mature, phenolic compounds polymerize, resulting in a decrease in the protein precipitating capacity (Makkar et al., 1988).
(d) Individual variation. The levels of anti-nutritive factors vary considerably between different parts of the same plant and there can be wide variation between individual trees of the same species (Josh) et m., 1985). This is true even of trees grown at the same site. With reports from different sites, it is not known to what extent observed variations are due to either environment or heredity.
In addition to the deleterious effects noted above, condensed tannins are generally, but not universally, considered to have two beneficial effects in ruminant nutrition. Firstly, they may protect labile plant proteins from microbial degradation in the rumen, thereby increasing the supply of high quality protein entering the duodenum and becoming available for absorption by the animal. Secondly, tannins appear to be implicated in the prevention of bloat in sheep and cattle by hindering the formation of stable protein foams in the rumen.
Since tannins occur in many valuable sources of animal feed, including forage crops, agricultural wastes and by-products, and tree foliage, it has been suggested that the digestibility of feeds could be improved by breeding programmes to develop low tannin crop lines. Given the protective function of tannins in the plant, however, such a procedure could lead to a loss of resistance to pests and diseases, thereby replacing a nutritional problem with an agronomic one (D'Mello, 1982).
Other approaches to the problem of tannins include water or alkali treatment or addition of adsorbents to remove tannins, or reaction with formalin to convert them to nonreactive resins. These processes all increase the cost of the feed and some lead to substantial losses of dry matter. As far as ruminants are concerned, it seems likely that the best approach would be to develop feeding systems to optimize the use of high tannin fodder in conjunction with other feed sources.
Toxic amino acids
Free, non-protein amino acids have been detected, principally in the leaves and seeds of many plant species, including a number of grain, pasture and browse legumes. They are capable of interfering with the normal metabolic processes of animals as a result of their effects on either amino acid biosynthesis and transport, or on activation and incorporation of amino acids into protein molecules (Hammond, 1987). They influence the rate of protein synthesis, cause the formation of aberrant proteins and affect RNA and DNA metabolism. Manifestations of toxicity range from simple reductions of food and nutrient utilization to lethal neurological disorders (D'Mello, 1992). The best known of these compounds are mimosine and canavanine. Mimosine is found at high concentrations in Leucaena and Mimosa spp. where it acts as a natural fungicide. Canavanine occurs in some tropical species of Canavalia and Sesbania, as well as in the temperate legumes Medicago sativa (lucerne) and Trifolium spp. (clovers), where it appears to have insecticidal properties.
Mimosine is typically found in foliage of mature Leucaena leucocephala at concentrations in the range of 2-5% (Brewbaker and Hylin, 1965), with higher levels of up to 10% in young, expanding leaves (Skerman et al., 1988). There is considerable variation in mimosine content between species of Leucaena and much attention has been paid to the production of interspecific hybrids, often of L. Ieucocephala with either L. diversifolia, L. pallida or L. pulverulenta, to combine low levels of mimosine with high forage production (e.g. Gupta et al., 1991). This approach could lead to reduced resistance to plant pests (D'Mello, 1982) and so may be inappropriate for general use.
It has long been recognized that Leucaena could cause depilation, growth reduction, general ill-health and reproductive disorders when fed to non ruminants at levels in the range of 5-15% of the diet on a dry matter basis (Hutton and Gray, 1957). Ruminants show tolerance to higher dietary levels but for the sake of safety, it has frequently been suggested that cattle should not consume more than 30% of their diet as Leucaena (Jones and Hegarty, 1984).
In non-ruminants, mimosine is absorbed directly into the blood stream where it acts as a protease inhibitor (Liener, 1980). In ruminants, however, rumen bacteria convert the mimosine to 3,4-dihydroxypyridine (DHP) which is then absorbed, acting as a goitrogenic agent to reduce the production of thyroxine.
In areas of the world such as Indonesia and Hawaii, where Leucaena is naturalized and animals have been fed it for many years, rumen bacteria have evolved which are capable of degrading DHP to nontoxic metabolites (Jones and Megarrity, 1983). In such areas, ruminants can be maintained on diets of 100% Leucaena for months at a time without showing symptoms of toxicity (Quintyne, 1987). The capacity to degrade DHP was transferred from goats in Hawaii to other goats and subsequently to cattle in Australia by inoculation with imported rumen liquor (Jones, 1985). Under normal grazing conditions, the rumen bacteria are able to move readily from one animal to another (Hammond, 1987), resulting in the rapid transfer throughout the herd, of the ability to utilize high levels of potentially toxic Leucaena. For a modest cost, ruminant resistance to mimosine toxicity can be introduced into areas where it does not occur naturally.
In Jamaica, a patented, anaerobic fermentation process has been developed which detoxifies ground Leucacna leaf meal for use in rations for nonruminants (Lewis, 1987). The process produces three major fractions:
· fermented leaf meal with negligible mimosine and DHP content, which can be used at high levels in diets for pigs and poultry;
· fuel gas, containing about 70% of methane and hydrogen; and
· liquid organic fertilizer with an approximate N:P:K ratio of 13:4:35 on a dry basis.
This process offers a useful technique to utilize the productive potential of Leucacna forage for feeding to non-ruminants. Canavanine is a water-soluble, heat-resistant structural analogue of arginine and as such is capable of radical interference in the metabolic pathways of livestock. Mammals are considered to be more tolerant of canavanine than are birds and the productive potential of C. ensiformis could be utilized to feed pigs, provided that the meal is subjected to heat treatment to deactivate the lectins that are also present in the beans (D'Mello, 1992). Cattle consuming more than 30% of their diet as seed or meal of Canavalia ensiformis are at risk (Skerman et al., 1988). Symptoms of toxicity include a low body temperature, a clear nasal discharge, frequent passing of clear urine, lameness, prostration and death. Skerman et al., (1988) recommended heat treatment to remove the toxicity, although Parra et al., (1988) indicated only a 50% reduction of canavanine content as a result of autoclaving. Treatment for 10 days with either 6% urea or 3% urea and 3% sodium hydroxide, led to complete elimination of the amino acid. The canavanine levels of beans of six cultivars of C. ensiformis ranged from 2.73-5.37%, suggesting that there may be scope for breeding to produce less toxic lines (Parra et al., 1988), although this may reduce plant pest resistance (D'Mello, 1992).
Cyanogenic glycosides
Hydrogen cyanide (HCN) is potentially the most serious anti-nutritional factor in fodder trees, since rapid absorption of the toxin can lead to the death of the animal. Symptoms of HCN poisoning are due to oxygen starvation at the cellular level and include laboured breathing (dyspnoea), intense red conjunctive, frothing at the mouth, bloat, convulsions and a staggering gait. Post-mortem examination often reveals a characteristic smell of almonds from the stomach contents.
Cyanogenic substrates, usually glycosides, react in the digestive tract of the animal with a hydrolyzing enzyme, often also found in the plant, to produce HCN. When HCN production is low, little threat is posed to the animal. Maslin et al., (1987) suggested that plants producing HCN at levels above 20 mg/100g (7.5 µmol/g) fresh weight could be considered as dangerous to livestock. The stomach contents tend to buffer the absorption of cyanide in ruminants (Goodchild and McMeniman, 1987), possibly due to its reaction with sugars or with sulphur compounds, to form harmless cyanhydrins or sulphocyanides respectively (Steyn and Rimington, 1935). Plants containing both large amounts of cyanogenic glycosides and an endogenous hydrolytic enzyme have the highest potential for toxicity (Maslin et al., 1987), although broad spectrum enzymes present in other fractions of the diet could react with the cyanogenic compounds to release HCN. In South African Acacia spp., leaves and immature pods produced higher levels of HCN than mature pods (Steyn and Rimington, 1935). A. robusta contained the glycoside but not the hydrolyzing enzyme and so was considered to be less dangerous than other species. HCN poisoning usually occurs only in sporadic outbreaks. A number of factors appear to be implicated, including the species and variety of browse consumed, the rate of ingestion and the availability of alternative feeds. It is more likely to occur during periods of drought or feed scarcity, when hungry animals consume large amounts of feed over a short period of time. Practical control measures can be taken to avoid cyanide poisoning:
· restrict the access to potentially poisonous plants and limit the grazing time during periods of feed scarcity;
· avoid feeding pods that are wet. Physically separate potentially dangerous feeds from water sources, since cold water appears to encourage the release of HCN;
· mix potentially toxic feeds with sulphur or molasses, or feed them in conjunction with licks containing these substances.
Alkaloids
The alkaloids are a heterogeneous group, with diverse chemical structures and this leads to problems of definition of both the group and their effects on livestock. Essentially they are complex compounds containing nitrogen, usually in heterocyclic and/or aromatic ring structures. They are basic in reaction, forming salts in the presence of acids. They are generally insoluble in water but extractable in organic solvents. Often they are poisonous to livestock but there are many examples of alkaloids with medicinal properties. Many are derived from amino acids and they have been described under headings that correspond to the amino acid unit (Southon and Buckingham, 1989). They are almost universally bitter in taste and while their role in plants has been variously ascribed, for example, as possible mechanisms of defence against plant pests (Levin, 1976), they are commonly held to represent evolutionary aberrations of nitrogen metabolism (Kingsbury, 1964).
Alkaloids are found in some 5-10% of all plants, being more common in tropical than in temperate species. Plant families such as Leguminosae, Amaryllidaceae and Compositae are noted for high levels of these compounds. Kingsbury (1964) indicated that more than 5000 alkaloids had been named and at least partially characterized. The alkaloid content of a plant is usually a feature of the cultivar, varying little with ecological factors such as climate, season and availability of water, although there can be wide differences between varieties of the same species. Unlike some other anti-nutritive factors such as tannins and amino acids which tend to be concentrated in certain plant parts, alkaloids are often uniformly distributed throughout the plant, all parts being equally dangerous (or beneficial) to livestock. The reaction of a given alkaloid in a given organism is usually quite specific, although there may be considerable variation between different alkaloids in the same animal species, or with the same alkaloid in different animals (Kingsbury, 1964).
One important subgroup in this category is that known as the pyrrolizidine alkaloids. These have a world-wide distribution, being present in both Leguminosae and Compositae (Smith and Culvenor, 1981). They are hepatotoxins which will cause fatal liver failure if ingested in sufficient quantities over a period of time (Molyneux and Ralphs, 1992). There is, however, a range of susceptibility to these compounds. Sheep are able to graze pastures considered dangerous to cattle, because of either better detoxification of the alkaloid in the rumen or through differences in hepatic activity (Hooper, 1978).
Clearly, shrub and tree species which contain appreciable quantities of alkaloids should be treated with caution where it is intended to utilize them as browse or fodder and they should be evaluated with a range of animal species before being recommended for general use. Nevertheless, the literature is not without its contradictions where alkaloids are concerned. Erythrina, a genus noted as a source of alkaloids, is attracting attention in Africa and South America as a multi-purpose tree suitable for use as fodder, and also for windbreaks, shade, fences and fuel (Brewbaker, 1989). Insufficient information is available to draw general conclusions regarding the effects of these compounds on animal production.
Differences in acceplability
There are many contradictions in the literature regarding the acceptibility of fodder from trees and shrubs, which may, or may not be of practical importance in commercial animal production. Reasons for the reported differences may be physical, chemical, or a combination of both. Some possible explanations are noted below.
(a) Acceptability can change during the year under the influence of season. Rodriguez et al., (1987) showed that with milking goats, consumption of dry matter from fresh, young Gliricidia foliage was less than with older, mature leaves. As the growing season progresses, the proportion of mature foliage on the tree will increase which should lead to improved utilization by goats.
(b) In some cases, it may take several days for animals to accept a new feed but once accustomed to it, they may then consume it readily (e.g. Atta-Krah and Sumberg, 1987, for Gliricidia).
(c) Preference for one feed over another does not necessarily imply poor acceptability of the latter when the former is absent. Observations from 'cafeteria' type trials where animals are given simultaneous access to a range of fodders, must be interpreted with care, since they reflect preferences rather than absolute acceptance or rejection, and then only at the time of the trial. A species which is rejected in favour of another may be well accepted if it is offered as the sole feed, or at a different time of the year.
(d) Within a single species, differences in acceptability can exist between varieties, accessions or provenances, individual trees, and even between parts of the same tree. There is much scope through selection and possibly even breeding to enhance the desirable characteristics of woody plants (e.g. Habit et al., 1981 for Prosopis tamarugo) and make them more available as fodder species.
(e) Within a single accession, acceptability can be influenced by edaphic and climatic conditions at the production site. The acceptability by animals of the same varieties of shrubby Stylosanthes spp. in Australia varies greatly between the sandy, infertile soils of the Cape York Peninsula and the more fertile soils of the Katherine region of the Northern Territory (W. Winter, Pers. Comm.)
References and further reading
ALLEN, O.N. and ALLEN, E.K. (1981) The Leguminosae: A Source Book of Characteristics, Uses and Nodulation. London: MacMillan Scientific and Medical Division.
AHN, J.H., ROBERTSON, B.M., ELLIOT, R., GUTTERIDGE, R.C. and FORD, C.W. (1989) Quality assessment of tropical browse legumes: tannin content and protein degradation. Animal Feed Science and Technology, 27:147-156.
ATTA-KRAH, A.N. and SUMBERG, J.E. (1987) Studies with Gliricidia septum for crop-livestock production systems in West Africa. In: Gliricidia septum (Jacq.) Walp: Management and Improvement. Proceedings, Workshop held at CATIE, Turrialba, Costa Rica, June 1987. Special Pub. 87-01. pp31-43. Hawaii, USA: Nitrogen Fixing Tree Association.
BATE-SMITH, E.C. (1962) The phenolic constituents of plants and their taxonomic significance. I Dicotyledons. Journal of the Linnean Society (Botany) 58: 95-173.
BATE-SMITH, E.C. (1973) Haemanalysis of tannins: the concept of relative astringency. Phytochemistry, 12: 907-912.
BREWBAKER, J.L. (1989) Nitrogen-fixing trees for fodder and browse in Africa. In: Alley Farming in the Humid Tropics. KANG, B.T. and REYNOLDS, L. (eds.) Proceedings, International Workshop held at Ibadan, Nigeria, March 1986. pp 55-70. Ottowa, Canada: International Development Research Centre.
BREWBAKER, J.L. and HYLIN, J.W. (1965) Variations in mimosine content among Leucaena species and related Mimosaceae. Crop Science, 5(4): 348-349.
D'MELLO, J.P.F. (1982) Toxic factors in tropical legumes. World Review of Animal Production, 18(4): 4146.
D'MELLO, J.P.F. (1992) Chemical constraints to the use of tropical legumes in animal nutrition. Animal Feed Science and Technology, 38: 237-261. FORAN, B.D. (1984) Central arid woodlands. In: Management of Australia's Rangelands. HARRINGTON, G.N., WILSON, A.D. and YOUNG, M.D. (eds). pp 299-315. Canberra, Australia: Commonwealth Industrial and Scientific Research Organization.
FORBES, M. (1986) The Voluntary Intake of Farm Animals. Stoneham, USA: Butterworths.
FOX, J.E.D. (1987) Potential of Australian Acacias from arid and semi-arid zones. In: Australian Acacias in 18 Develop�ng Countries. TURNBULL, J.W. (ed). Proceedings, International Workshop held at Gympie, Australia, August 1986. ACIAR Proceedings No.16. pp 17-28. Canberra, Australia: ACIAR.
GOODCHILD, A.V. and McMENIMAN, N.P. (1987) Nutritive value of Acacia foliage and pods for animal feeding. In: Australian Acacias in Developing Countries. TURNBULL, J.W. (ed.) Proceedings, International Workshop held at Gympie, Australia, August 1896. ACIAR Proceedings No.16. ppl 01-106. Canberra, Australia: ACIAR.
GUPTA, B.K., AHUJA, A.K. and KHEJURIA, H.N. (1991) Fodder analysis of new Leucaena varieties. Leucaena Research Reports, 12: 40-41.
HABIT, M.A., CONTRERAS, D.T. and GONZALEZ, R.H. (1981) Prosopis tamarugo: fodder tree for the arid zones. FAO Plant Production and Protection Paper No.25. Rome: Food and Agricultural Organization.
HAGERMAN, A.E. and BUTLER, L.G. (1981) The specificity of proanthocyanidin-protein interactions. Journal of Biological Chemistry. 256: 4494-4497.
HAGERMAN, A.E. and BUTLER, L.G. (1989) Choosing appropriate methods and standards for assaying tannin. Journal of Chemical Ecology, 15(�): 1795-1810.
HAMMOND, A.C. (1987) Chemical, anatomical and other antiquality factors limiting forage utilization. In: Forage Livestock Research Needs for the Caribbean Basin. MOORE, J.E., QUESENBERRY, K. H. and MICHAUD, M.W. (eds) Proceedings, Workshop held at Tampa, USA. February, 1987. pp 59-69. Gainsville, USA: Caribbean Basin Advisory Group/University of Florida.
HOOPER, P.T. (1978) Pyrrolizidine alkaloid poisoning: pathology with particular reference to differences in animal and plant species. In: Effects of Poisonous Plants on Livestock. KEELER, R.F., VAN KAMPEN, K.R. and JAMES, L.F. (eds). New York: Academic Press.
HUTTON, E.M. and GRAY, S.G. (1957) Problems in adapting Leucaena glauca as a forage for the Australian tropics. Empire Journal of Experimental Agriculture, 27: 187-196.
JONES, R.J. (1985) Leucaena toxicity and ruminal degradation of mimosine. In: Plant Toxicology. SEAWRIGHT, A.A., HEGARTY, M.P., JAMES, L.F. and KEELER, R.F. (eds.) Proceedings, Australia-USA Poisonous Plants Symposium, held at Brisbane, Australia, May 1984. pp. 111-119. Yeerongpilly, Australia: Queensland Poisonous Plants Committee.
JONES, R.J. and HEGARTY, M.P. (1984) The effect of different proportions of Leucacna in the diet of cattle on the growth, feed intake, thyroid function and secretion of DHP. Australian Journal of Agricultural Research, 35(2): 317-325.
JONES, R.J. and MEGARRITY, R.G. (1983) Comparitive toxicity responses of goats fed on Leucaena leucocephala in Australia and Hawaii. Australian Journal of Agricultural Research, 35: 781-790.
JOSHI, U.N., ARORA, S.K., PARODA, R.S., JATASARA, D.S. and RANA, D.S. (1985) Chemical composition of Prosopis leaves to show the presence of anti-nutritive factors responsible for low in vitro dry matter digestibility. Nitrogen Fixing Tree Research Reports, 3: 20-21.
KINGSBURY, J.M. (1964) Poisonous Plants of the United States and Canada. Englewood Cliffs, USA: Prentice Hall Inc.
KUMAR, R. and SINGH, M. (1984) Tannins: their adverse role in ruminant nutrition. Journal of Agricultural Food Chemistry, 32(3): 447-453.
KUMAR, R. and VAITHIYANATHAN, S. (1990) Occurrence, nutritional significance and effect on animal productivity of tannins in tree leaves. Animal Feed Science and Technology, 30: 21-38.
LEVIN, D.A. (1976) Alkaloid-bearing plants: an ecogeographic perspective. American Naturalist, 110: 261-274.
LEVIN, D.A. and YORK, B.M. (1978) The toxicity of plant alkaloids: an ecogeographic perspective. Biochemical Systems and Ecology, 6: 61-76.
LEWIS, C.E. (1987) (ed.) Proceedings, First Caribbean Leucaena Industries Symposium, held at Kingston, Jamaica, August, 1987. Kingston, Jamaica: Enerplan Ltd.
LIENER, I.E. (1980) Toxic Constituents of Plant Foodstuffs. London: Academic Press.
MAKKAR, H.P.S., DAWRA, R.K. and SINGH, B. (1988) Changes in tannin content, polymerization and protein precipitation capacity in oak (Quercus incana) leaves with maturity. Journal of Science of Food and Agriculture, 44: 301-307.
MANGAN, J.L. (1988) Nutritional effects of tannins in animal feeds. Nutrition Research Reviews, 1: 209-231.
MASLIN. B.R., CONN, E.E. and DUNN, J.E. (1987) Cyanogenic Australian species of Acacia: a preliminary account of their toxicity potential. In: Australian Acacias in Developing Countries. TURNBULL, J.W. (ed) Proceedings, International Workshop held at Gympie, Australia, August 1986. ACIAR Proceedings No.16. pp 107-111. Canberra, Australia: ACIAR.
McLEOD, M.N. (1974) Plant tannins - their role in forage quality. Nutrition Abstracts and Reviews, 44: 803-815.
McNAUGHTON, S.J. (1987) Adaptation of herbivores to seasonal changes in nutrient supply. ln: Nutrition of Herbivores. HACKER, J.B. and TERNMOUTH, J.H. (eds.) pp391-408. Sydney, Australia: Academic Press.
MOLYNEUX, R.J. and RALPHS, M.H. (1992) Plant toxins and palatability to herbivores. Journal of Range Management, 45: 13-18.
PARRA, R., ESCOBAR, A., de VIERMA, C., VARGAS, R., VIERA, J., DOMINGUEZ, M.G. and MORA, M. 1988. The potential and constraints to Canavalia spp. as an animal feed resource. In: Forage Legumes and Other Local Protein Sources as Subsitutes for Imported Protein Meals. WALMSLEY, D (ed.) Proceedings CTA/CARDI Seminar, Kingston, Jamaica, December 1987. pp. 154-180. Port of Spain, Trinidad: CARDI.
QUINTYNE, R.C. (1987) Utilization of Leucaena leucocephala as a livestock feed in Barbados. In: First Caribbean Leucaena Industries Symposium. LEWIS, C.E. (ed.) Proceedings, Workshop held at Kingston, Jamaica. pp. 10-14. Kingston, Jamaica: Enerplan Ltd.
RODRIGUEZ, Z., BENAVIDES, J.E., CHAVES, C. and SANCHEZ, G. (1987) Produccion de leche de cabras estabu lad as a limentadas con foliaje de madero negro (Gliricidia septum) y de poro (Erythrina poeppigiana) y suplementadas con platano pelipita (Musa sp. cv. Pelipita) In: Gliricidia septum (Jacq.) Walp: Management and Improvement. Proceedings, Workshop held at CATIE, Turrialba, Costa Rica, June 1987. Special Pub. 87-01. pp 212-216. Hawaii, USA: Nitrogen Fixing Tree Association.
SKERMAN, P.J., CAMERON, D.G. and RIVEROS, F. (1988) Tropical Forage Legumes. 2nd edn. Rome: Food and Agriculture Organization.
SMITH, L.W. and CULVENOR, C.C.J. (1981) Plant sources of hepatotoxic pyrrolizidine alkaloids. Journal of Natural Products, 44(1): 129-152.
SOUTHON, I.W. and BUCKINGHAM, J. (1989) Dictionary of Alkaloids. London, UK: Chapman and Hall. 2 vols.
STEYN, D.G. and RIMINGTON, C. (1935) The occurrence of cyanogenetic glycosides in South African species of Acacia. Onderstepoort Journal of Veterinary Science and Animal Industry, 4(1): 51-73.
TANNER, J.C., REED, J.D. and OWEN, E. (1990) The nutritive value of fruits (pods with seeds) from four Acacia spp. compared with extracted noug (Guizotia abyssinica) meal as supplements to maize stover for Ethiopian highland sheep. Animal Production, 51: 127133.
VAN HOVEN, W. (1985) The tree's secret weapon. South African Panorama, 30(3): 34-37.
Other titles in the Use of Trees by Livestock series include:
1 Prosopis
2 Acacia
3 Gliricidia
5
Quercus
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Use of Trees by Livestock (NRI, 1994, 160 p.)
Use of Trees by Livestock: Quercus
R.T. Paterson
Acknowledgements
© Crown copyright 1993
The Natural Resources Institute (NRI) is an internationally recognized centre of expertise on the natural resources sector in developing countries. It forms an integral part of the British Government's overseas aid programme. Its principal aim is to alleviate poverty and hardship in developing countries by increasing the productivity of their renewable natural resources. NRI's main fields of expertise are resource assessment and farming systems, integrated pest management, food science and crop utilization.
NRI carries out research and surveys; develops pilot-scale plant, machinery and processes; identifies, prepares, manages and executes projects; provides advice and training; and publishes scientific and development material.
Short extracts of material from this publication may be reproduced in any non-advertising, nonprofit-making context provided that the source is acknowledged as follows:
Paterson, R.T. (1993) Use of Trees by Livestock 5: Quercus. Chatham, UK: Natural Resources Institute.
Permission for commercial reproduction should be sought from:
The Head, Publishing and Publicity Services, Natural Resources Institute, Central Avenue, Chatham Maritime, Kent, ME4 4TB, United Kingdom.
No charge is made for single copies of this publication sent to the governmental and educational establishments, research institutions and non-profitmaking organizations working in countries eligible for British Government Aid. Free copies cannot normally be addressed to individuals by name but only under their official titles. Please quote LC5 when ordering.
Natural Resources Institute
ISBN: 0 85954 365-X
Price
£2.00
This booklet is printed on paper produced using the residues from sugar-cane processing.
Foreword
The importance of trees and shrubs in the feeding of animals in the tropics and sub-tropics has long been recognized by livestock owners. In arid areas where the growth of herbaceous plants is limited by lack of moisture, leaves and edible twigs of trees and shrubs can constitute well over 50% of the biomass production of rangeland. At high altitudes, tree foliage may provide over 50% of the feed available to ruminants in the dry season, branches being harvested and carried to the animals. Even in regions of higher rainfall where grass supplies the major proportion of the dry matter eaten by ruminants, tree leaves and fruits can form an important constituent of the diet, particularly for small ruminants.
In the last two decades interest in the planting of trees as a source of feed for livestock has been encouraged by workers in research and development, but in contrast to the hundreds of indigenous species which are used as fodder, attention has focussed on a limited number of introduced species. Thus there are many publications reporting the chemical composition of Leucaena leucocephala leaves and suggesting management strategies for utilization of the tree for fodder, but it is more difficult to find information on alternative genera which might be equally, or more, appropriate.
The aim of this series of publications is to bring together published information on selected genera of trees which have the potential to increase the supply of fodder for ruminants. Each booklet summarizes published information on the fodder characteristics and nutritive value of one genus, with recommendations on management strategies, where available. Further, since the leaves of woody species frequently contain secondary compounds which may have an antinutritional, or toxic, effect, a separate booklet summarizes the effects of a number of these compounds. It is hoped that the booklets will provide useful resource material for students, research and extension workers, interested in promoting the use of trees as a source of fodder for ruminants.
Further copies of this booklet or others in the series can be obtained by writing to Publishing and Publicity Services at the Natural Resources Institute.
Margaret Gill
Livestock Production
Programme
Genus Quercus
Family FAGACEAE
Principal species Quercus alba
Q.
calliprinos
Q. coccipra
Q. falcata
Q. gambelii
Q. glauca
Q.
grisea
Q. havardii
Q. flex
Q. incana
Q. lamellosa
Q.
macrocarpa
Q. pubescens
Q. robur (syn. Q. pedunculata)
Q. rubra (syn.
Q. borealis)
Q. semecarpifolia
Q. suber
Q. velutina (syn. Q. tinctoria)
Common name Oak
Summary
The leaves and acorns of many oak species are avidly eaten by cattle, sheep, goats, horses and pigs. Oak fodder contains relatively high levels of polyphenolic compounds and is not highly digestible but it is often available at times of the year when other fodder is in short supply. Its crude protein content is reasonable and it can be a useful supplement to poor-quality grazing or to cereal straw-based diets.
Oak toxicity occurs sporadically when animals consume too high a proportion of either leaves or acorns in their diets. Symptoms of toxicity appear within a few days and death follows rapidly. The provision of alternative feed, sometimes together with supplements of calcium reduce the incidence of poisoning.
Oak trees are used for many purposes including timber for tools, construction, ship-building and joinery. The wood is used for fuel and makes good charcoal. The bark provides tannins, inks and dyes for fabrics and leather work and cork for use in bottling, linoleum products etc. Acorns are sometimes eaten by humans and the leaves are used to feed silkworms and cochineal insects. They are important multi-purpose trees in many parts of the world.
Description and distribution
Quercus is a genus of some 600 woody, perennial species of diverse habit and leaf shape, ranging in size from shrubs to tall trees. Oaks are characterized by their fruit which takes the form of an acorn (Kingsbury, 1964), developing from a tricarpellate, inferior ovary and subtended by a cup formed by numerous overlapping scale leaves. The acorns mature over either one (white oaks) or two (black oaks) years. Trees may be either evergreen or deciduous but where they lose their leaves in autumn, they produce well developed winter buds.
The genus has a wide geographical distribution. In the Americas, it is found from the far north down into the tropical western parts of South America. It is also native to temperate and tropical Eurasia and both north and south Africa. Prior to widespread felling for agricultural purposes, oaks were a major component of many European forests. There appears to be natural hybridization, at least between some European species, and synonymy seems to have resulted from imperfect genetic isolation of species (Mabberly, 1987).
Many species have economic uses based either on their strong, durable, close-grained timber or on the bark which provides cork and a range of extracts used for inks, dyes, leather tanning and medicinal purposes (Uphof, 1968; Willis, 1973). The acorns are commonly used as food for both humans and livestock and the leaves are fed to domestic animals, including silkworms (Uphof, 1968). Nevertheless, oak toxicity has been described in a range of animals, including cattle, buffaloes, horses, sheep, pigs, rabbits and dogs in a number of countries (Boughton and Hardy, 1936, in USA; Clarke and Cotchin, 1956, in UK; Kingsbury, 1964, in France, Germany and New Zealand; Neser et al., 1982, in South Africa; Shi, 1988, in China).
Fodder characteristics
The plant parts of oaks and the season in which they are most readily consumed by livestock both depend to a large extent on the growth habit of the particular species under consideration. In the southwest of North America where shrubby species such as Q. grisea, Q. gambelii, and Q. dumosa are common, leaves, buds and twigs are browsed avidly by goats and to a lesser extent by sheep and cattle (Ahmed, 1982; Nunez-Hernandez et al., 1989; Holechek et al., 1990). In southern Europe, this is also true of Q. flex and regrowths of Q. pubescens (Guerro and Boza, 1983; Meuret et al., 1985; Meuret, 1988), while in Israel, leaves and young stems of Q. calliprinos are an important source of feed for cattle and goats (Perevolotsky et al., 1993). The reported levels of voluntary intake of native oak browse by goats in France, Greece and Israel are higher than with the common oak species in North America at equivalent levels of digestibility and protein content. This may be attributable to the evolution of adaptive digestion behaviour over a much longer period in the Old World than has so far been possible in North America (Perevolotsky et al., 1993). In India and Nepal leaves of a number of species such as Q. incana, Q. semecarpifolia and Q. glauca are harvested either for direct feeding of fresh material or for storage for use in times of fodder shortage (Lohan et al., 1983; Bajracharya et al., 1985). In such situations, the importance of acorns in animal nutrition is limited and oaks make their greatest contribution in the spring and summer.
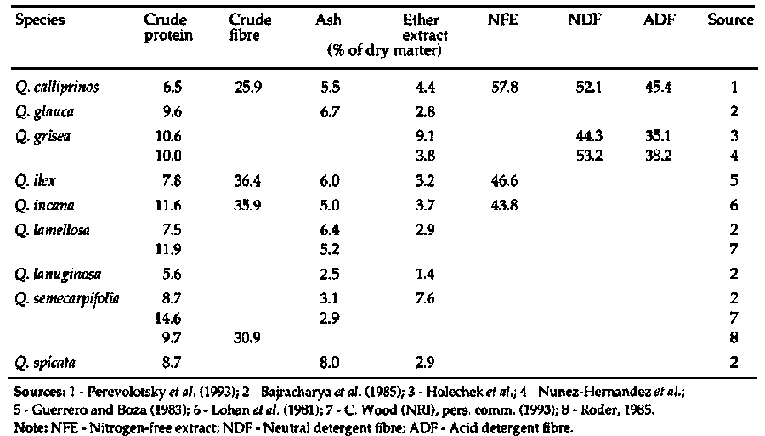
Table 1 - Analysis of edible leaves
and twings of Quercus spp.
In contrast to this, acorns from tall, mature trees of species such as Q. robur and Q. alba constitute the major source of oak fodder in the northeastern part of North America (Kingsbury, 1964), northern Europe (Clarke and Cotchin, 1956) and South Africa (Neser et al., 1982). Warm, dry summers tend to favour heavy crops of acorns. Autumn storms and winter snows can then lead to the simultaneous shedding of large quantities of fruits which can become an important source of feed when grazing and other browse may be unattractive, unavailable or scarce (Clarke and Cotchin, 1956; Holliman, 1985; Spier et al., 1987) late in the year. In some areas, fallen leaves are also consumed from the litter on the ground under oak trees, but this material is not particularly attractive, being of poor quality and low digestibility (Forwood and Owensby, 1985).
Some typical data for the chemical analysis of leaves, buds and edible twigs of a number of oak species are shown in Table 1. On the basis of these data, Quercus spp. could be considered as relatively good sources of fodder, since in comparison with other commonly available browse species they show reasonably high levels of crude protein, moderate amounts of crude fibre and ether extract (fats) and low residues of ash (Bajracharya et al., 1985).
In vitro dry matter digestibility (IVDMD) averaged only 36.9% from mid-September to late October for Q. macrocarpa, and was only slightly better for a Q. macrocarpa hybrid (similar to Q. alba) with 42.6% (Forward and Owensby, 1985).
In general, however, in vitro analyses appeared to underestimate the digestibility of oak fodder when it was below about 60%, the discrepancy becoming greater at lower levels (Perevolotsky et al., 1993). In vivo digestibility of a range of oak species from both Europe and North America appeared to fall in the range of 47-49% when measured in goats, although this could be increased by some 4-9% by the inclusion in the diet of high-quality meals made from lucerne (Medicago saliva) or soyabeans (Meuret, 1988; Nastis and Malechek, 1988). When fed as the sole dietary component to pregnant adult goats, Q. calliprinos was slightly deficient in protein, but provided adequate energy for maintenance (Perevolotsky et al., 1993)
The inclusion of small proportions of Q. gambelii and Q. grisea in goat rations based on either lucerne or barley straw resulted in significant reductions of in vivo digestibility of the overall rations (Nastis and Malechek, 1981; Holechek et al., 1990). This effect may be the result of the presence of anti-nutrive factors in the oak fodder.
It has been noted (Nastis and Malechek, 1981) that where oak browse constitutes a large proportion of the ration, its low level of digestibility may reduce voluntary intake. This, in turn, may result in a diet which is deficient in metabolizable energy, especially if consumed together with native forage of poor nutritional quality. When used as a supplement at levels of intake below 25% of the diet, however, Q. incana is capable of increasing the digestibility and voluntary intake of wheat straw (Lohan et al., 1983). It would appear that oak foliage is best used at relatively low levels of intake to supplement other, poor quality feedstuffs.
While not a common practice, the use of oak sawdust has been studied as the roughage component of diets for beef cattle on high-energy finishing rations (el-Sabban et al., 1971). Sawdust could be successfully used at levels of up to 15% as fed, of a ration based on shelled corn and soyabean meal, without detrimental effects on growth rates or carcass characteristics. Coarse sawdust produced from a circular saw (modulus of fineness 4.56) was preferable to fine sawdust from a band saw (modulus of fineness 2.40), as the coarser material appeared to improve rumen condition and reduce the incidence of liver abscesses.
Anti-nutritive factors
The poor digestibility of oak foliage is generally attributed to the high content of phenolics (including tannins) which is found in all species. Using the Folin-Denis reagent, Nastis and Malechek (1981) found ll.l% tannic acid equivalent of phenolic compounds in the immature leaves of Q. gambelii and 8.7% in the mature leaves, while Nunez-Hernandez et al., (1989) and Holechek et al. (1990) found 7.9% in the immature leaves of Q. grisea and 5.4% in the mature leaves. From a single tree of Q. robur, Feeny and Bostock (1968) found that the total tannin content increased from less than 1% in early spring to 5% in late summer, the increase being largely due to an increase in the level of condensed tannin. These authors concluded that the period of heaviest insect attack (early spring) coincided with the lowest levels of total tannins and the near complete absence of condensed tannins in the leaves of Q. robur thus illustrating the protective role of tannins. In contrast, total phenolics were found to be highest in young leaves of Q. gambelii (Nastis and Malechek, 1981) and Q. havardii (Dollahite et al., 1966), the content falling with leaf maturity. In Q. incana the total phenolic content and protein precipitating capacity decreases with advancing leaf maturity while condensed tannins increase (Makkar et al., 1988).
The total phenolic content of acorns varies with both the species of oak and the stage of maturity of the fruit (Basden and Dalvi, 1987). When extracted with Folin-Ciocalteu reagent, Q. alba (0.41-2.54%) contained less phenolics (tannic acid equivalent) than Q. rubra (3.72-4.47%) or Q. velutina (3.29-6.13%) when expressed as a percentage of air-dry weight of acorns with the cups removed. Green acorns contained from 1.5 to 3 times the phenolic levels of ripe acorns.
Bowns (1988) noted that many toxic species within a range of potentially poisonous plants, including Q. gambelii and species of Prunus, Cercocarpus, Delphinium and Astragalus, are considered by farmers to be good, or even excellent, forages with only beneficial effects on livestock for much of the year. Under certain conditions, however, animals may consume excessive amounts of these plants too rapidly. They then suffer the effects of toxicity, resulting in reduced productivity, or even death.
Although oaks may sometimes appear to be only nutritionally beneficial to livestock, the potential toxicity of both acorns and foliage has long been recognized. Few early reports adequately identified the toxic species of Quercus, but the physical symptoms of poisoning do not appear to vary significantly with either tree species or part of the plant ingested. As early as the 19th century, poisoning was attributed to the high tannin content of oaks (see Kingsbury, 1964). Dollahite et al. (1962) failed to produce symptoms of oak toxicity by the oral administration of large amounts of tannic acid to cattle, sheep and goats but repeated doses of either commercial tannins or extracts of oak leaves produced symptoms in rabbits that approximated natural oak poisoning. While it is often assumed that toxicity is due to tannic acid, garlic acid and pyrogallol (Basden and Dalvi, 1987) or to other, unspecified, low molecular weight compounds produced by the breakdown of hydrolyzable tannins in the gut (Shi, 1988), Neser et al., (1982) suggested that the possibility of a mycotoxin should not be ignored. Moulds are often observed on fallen acorns and these could produce the toxic principal responsible for the poisoning of livestock. The exact cause of oak toxicity is still open to debate although the presence of potentially toxic compounds other than phenolics in oak leaves and acorns has not been reported.
The toxicity of oak is sporadic and reportedly low, even though buds and immature leaves are relished by cattle, sheep and goats. Harsh weather conditions including drought, high winds or heavy snow, are often associated with outbreaks of oak poisoning (Clarke and Cotchin, 1956; Neser et al., 1982; Spier et al., 1987). Such weather can cause the simultaneous shedding of large quantities of acorns at times when other forage is scarce. The large buds may be the only browse available in late winter or early spring and under these conditions animals may consume excessive amounts of oak fodder. Toxicity symptoms appear after a few days and rapidly become acute. Symptoms, which are relatively consistent across animal species, include anorexia, constipation, rough coat, dry muzzle, abdominal pain, excessive thirst and frequent urination. Ruminant animals often also suffer rumen stasis. If death does not occur, foetid, hemorrhagic diarrhoea sets in, accompanied by subcutaneous oedema in the ventral part of the body. The pulse becomes thin and rapid and there may be a brownish discharge from the nostrils. Severe hypocalcaemia and anaemia are noted and renal failure follows. High levels of blood urea can be measured and in both sheep and cattle: Holliman (1985) showed that animals with urea levels higher than 50 mmol/litre of blood rarely survived. Occasionally, death occurs within 24 hours of the onset of the symptoms but 3-10 days is more common.
Animals differ in their susceptibility to oak poisoning. In many countries acorns are collected for feeding to pigs as a supplement to their normal diets. Kingsbury (1964) speculated that the absence of negative consequences to this practice could be due either to the use of oak species with low toxic potential, the consumption of insufficient oak material to provoke toxicity, or to a greater tolerance of oak toxicity in pigs than in other animals. It is generally held that amongst domestic livestock, pigs and goats show greatest tolerance and cattle and buffaloes the greatest susceptibility. Sheep and horses are considered to be intermediate with regard to oak poisoning (Clark and Cotchin, 1956; Kingsbury, 1964; Neser et al., 1982; Holliman, 1985). Range cattle appear to be able to eat oak foliage at levels of up to half of their diet on a dry matter basis without suffering ill effects (Kingsbury, 1964), while both Angora and dairy goats can consume 80-85% of their dietary dry matter as browse of Q. gambelii (Davis et al., 1975; Nastis and Malechek, 1981), without showing either physical or physiological symptoms of toxicity.
In herds of cattle given free access to browse containing Quercus spp., there is a marked effect of age on susceptibility to poisoning. In the Ermelo region of South Africa, Neser et al. (1982) reported that in a herd of 80 animals ranging in age from one to eight years, 22 animals suffered from poisoning and of these, 16 died. All the affected animals were less than two years old. In the Lake District of UK, in an outbreak of toxicity in 1984, confirmed cases included cattle ranging in age from three months to adult cows, and sheep ranging from fattening lambs to adult ewes. The most susceptible animals were the young cattle (Holliman, 1985). In California, in an outbreak resulting in the death of 2700 cattle in 1985, Ostrowski et al. (1989) noted that the most affected animals were calves weighing 100 - 180 kg.
Management
In most parts of the world, little attempt is made to manage oak trees specifically for the provision of fodder to ruminant livestock. Acorns and senescent leaves are generally seen as seasonal feeds which are utilized opportunistically as they fall to the ground, while accessible buds are browsed in a similar manner as they open and attract the attention of animals. Under ranching conditions, the major challenge to management is to ensure, in times of stress when other feeds may be in short supply, that the animals do not consume a diet too rich in oak forage. Poisoning has been avoided by ad lib. provision of alternative feed to reduce the intake of fodder from oak trees. Dollahite et al. (1966) recommended the use of well-balanced supplementary feed to which calcium hydroxide had been added at a rate of 15% by weight (reducing to a minimum of 10% if the higher rate resulted in acceptability problems) to counteract the characteristic hypocalcaemia associated with oak toxicity. In contrast to this, Spier et al. (1987) reported that when unseasonal snow in California in April 1985 denied animals their normal forage and cattle ingested toxic quantities of buds of Q. douglesii, the immediate provision of hay, even in the absence of additional minerals, resulted in minimal stock losses. A total of some 2700 animals on 60 ranches died of oak toxicosis in the reported outbreak. Most of these received no supplementary feeding.
As noted above, cattle under two years of age appear to be at most risk from oak poisoning. It is therefore obvious that when conditions are such that excessive amounts of oak fodder may become available, young cattle should be removed to safe areas or provided with supplementary feed to reduce the risk of toxicity.
In Europe, it has been shown that leaves and twigs of Q. flex can be used as a protein supplement for goats (Guerrero and Boza, 1983; Meurat, 1988). The plant material can be browsed or harvested and fed fresh, and is particularly valuable during winter, or in times of drought.
In India and Nepal, oak trees are of particular importance at higher altitudes where the number of browse species is limited. In this region, it is common practice to harvest oak leaves as animal fodder during the growing season, either for immediate use or for storage on the farm for use during times of feed scarcity (Bajracharya et al., 1985; Makkar et al., 1986). It was originally thought that Indian oaks had no toxic effects on livestock (Chopra et al., 1955) but more recent work has shown a reduction in the productivity of cattle fed moderate levels of Q. incana and the manifestation of toxicity symptoms when fed at higher levels (Lohan et al., 1983; Makkar et al., 1986). Sole feeding of oak leaves to yearling calves produced toxic effects within two days and one animal out of a group of four died within four days (Negi et al., 1979). It would appear that the common perception of oak fodder is as a valuable resource to be used sparingly to supplement wheat straw or poor quality volunteer grasses during periods of feed shortages. This concept, resulting in the consumption of relatively low levels of oak leaves, would tend to maximize the efficient use of the scarce supplement and minimize the danger of stock losses due to poisoning.
Alternative uses
The many and varied economic uses of oak trees in their differing environments have been described by numerous authors including Smith (1882), Uphof (1968) and Mabberley (1987).
The wood of many species is heavy, close-grained, strong, durable and of a pleasing colour. It is used in numerous countries of the Americas, Europe and the Far East for construction, agricultural implements and small tools, carriages, boat-building, furniture and interior finishes, cooperage, basket-making, railway sleepers, fences etc. Species particularly valued for timber include Q. alba, Q. eduardi, Q. garryana, Q. flex, Q. macrocarpa, Q. pubescens, Q. robur, Q. rubra and Q. virginiana.
Several species, such as Q. bicolor, Q. grisea, Q. macrocarpa and Q. nigra are used for fuel, while Q. incana and Q. marylandica are reputed to make excellent charcoal.
In Japan, trunks of dead Q. glandulifera trees are used for the cultivation of edible mushrooms.
The bark of many oak species is utilized in industry. Q. suber, and occasionally other species such as Q. pseudosuber and Q. occidentalis, are cultivated for the production of cork, principally in Portugal and around the northern Mediterranean. Q. dendata, Q. lusitanica, Q. prinus, Q. robur and Q. velutina amongst others are used for tanning leather, while textile dyes are extracted from Q. tauricola, Q. variabilis and Q. velutina. Powdered bark of Q. alba, containing about 10% tannin, is used medicinally as an astringent and a tonic. North American Indians used a decoction of the bark of Q. Iyrata to treat dysentery and stomach ache.
Oak galls (commonly known as oak apples) are growths produced by perforations made by egglaying insects on leaves and twigs. These callouses, known commercially in relation to the area of origin as Aleppo, Levant, Mecca and Turkish galls, contain 36-58% tannin (Mabberly, 1987) and are collected for use in leather processing, the manufacture of inks and dyes and in ointments and suppositories. Q. infector�a produces the best known commercial galls but other species such as Q tauricola and Q robur are also exploited for these purposes.
The acorns of many species, including Q. cuspidata, Q. emoryi, Q. gambelii, Q. flex, Q glabra, Q. nigra and Q. persica are eaten, either raw or cooked, by humans in many parts of the world. They may be roasted, boiled or sometimes made into a flour for baking. Species such as Q. obtusiloba and Q. robur are used as coffee substitutes. Q. cerris is a source of oak manna (gaz or gazu much) used to make a sweet-meat in Iran.
The leaves of Q. obtusiloba have been used as cigarette papers by North American Indians, while silkworms (Antheroea pernyi) are fed on leaves of Q faberi and other oak species in China. Q. coccifera is used for feeding cochineal insects (Coccus ilicis).
Many oak species are handsome trees and much prized as ornamentals.
References and further reading
AHMED, A.E.T.S. (1982) Selection, intake and utilization of browse by goats. Dissertation Abstracts International, 42(8):3106.
BAJRACHARYA, D., BHATTARAI, T.B., DHAKAL, M.R., MANDAL, T.N., SHARMA, M.R., SITAULA, S. and VIMAL, B.K. (1985) Some feed values for fodder plants from Nepal. Vereinigungfur Angewandte Botanik, 59:357-365.
BASDEN, K.W. and DALVI, R.R. (1987) Determination of total phenolics in acorns from different species of oak trees in conjunction with acorn poisoning in cattle. Veterinary and Human Toxicology, 29(4):305-306.
BOUGHTON, I.B. and HARDY, W.T. (1936) Oak poisoning in range cattle and sheep. Journal of the American Veterinary Association, 89:157-162.
BOWNS, J.E. (1988) The importance of poisonous plants as forages in the intermountain region. In: The Ecology and Economic Impact of Poisonous Plants on Livestock Production. JAMES, L.F., RALPHS, M.H. and NIELSEN, D.B. (eds). Boulder, USA: Westview Press. pp. 377-390.
CHOPRA, R.N., BADHWAR, R.L. and GHOSH, S.C. (1955) Poisonous Plants of India. New Delhi: Indian Council of Agricultural Research. Part 1, p. 20. Part 2, pp. 832-833.
CLARKE, M.A. and COTCHIN, E. (1956) A note on the toxicity of the acorn. British Veterinary Journal, 112:135-139.
DAVIS, G.G., BARTEL, L.E. and COOK, C.W. (1975) Control of gambel oak sprouts by goats. Journal of Range Management, 28(3):216-218.
DOLLAHITE, J.W., HOUSEHOLDER, B.S. and CAMP, B.J. (1966) Effect of calcium hydroxide on the toxicity of post oak (Quercus stellata) in calves. Journal of the American Veterinary Medical Association, 148(8):908-912.
DOLLAHITE, J.W., PIGEON, R.F. and CAMP, B.J. (1962) The toxicity of garlic acid, pyrogallol, tannic acid and Quercus havardi in the rabbit. American Journal of Veterinary Research, 23:1264-1266.
FEENY, P.P. and BOSTOCK, H. (1968) Seasonal changes in the tannin content of oak leaves. Phytochemistry, 7:871-880.
FORWOOD, J.R. and OWENSBY, C.E. (1985) Nutritive value of tree leaves in the Kansas Flint Hills. Journal of Range Management, 38(1):61-64.
GUERRERO, J.E. and BOZA, J. (1983) El ramon de encina en la alimentaci�n del ganado cabr�o. Avances en Alimentaci�n y Mejora Animal, 24(8/9):287-289.
HOLECHEK, J.L., MUNSHIKPU, A.V., SAIWANA, L., NUNEZ-HERNANDEZ, G., VALDEZ, R., WALLACE, J.D. and CARDENAS, M. (1990) Influences of six shrub diets varying in phenol content on intake and nitrogen retention of goats. Tropical Grasslands, 24:93-98.
HOLLIMAN, A. (1985) Acorn poisoning in ruminants. The Veterinary Record, 116:546.
KINGSBURY, J.M. (1964) Poisonous Plants of the United States and Canada. Englewood Cliffs, USA: Prentice Hall Inc. pp. 444-446.
LOHAN, O.P., LALL, D., VAID, J. and NEGI, S.S. (1983) Utilization of oak tree (Quercus incana) fodder in cattle rations and fate of oak-leaf tannins in the ruminant system. Indian Journal of Animal Science, 53(10):1057-1063.
MABBERLY, D.J. (1987) The Plant Book. Cambridge, UK: Cambridge University Press.
MAKKAR, D.J., DAWRA, R.K. and SINGH, B. (1988) Changes in the tannin content, polymerisation and protein precipitation capacity in oak (Quercus incana) leaves with maturity. Journal of the Science of Food and Agriculture, 44:301-307.
MAKKAR, H.P.S., SINGH, B. and DAWRA, R.K. (1986) Can oak leaves be fed to animals? Farmers and Parliament, 24:17-18.
MEURET, M. (1988) Digestibilit� du feuillage de ch�ne vert (Quercus flex) distribu� frais � des caprins entraines au p�turage sur parcours. Reproduction, Nutrition, Developpement, 28(1):89-90.
MEURET, M., BARTIAUX-THILL, N. and BOURBOUZE, A. (1985) Evaluation de la consommation d'un troupe de ch�vres laiti�res sur parcours forestier: methode d'observation directe des coups de dents; methode du marqueur oxyde de chrome. Annales de Zootechnie, 34(2):159-179.
NASTIS, A.S. and MALECHEK, J.C. (1981) Digestion and utilization of nutrients in oak browse by goats. Journal of Animal Science, 53(2):283-290.
NASTIS, A.S. and MALCHEK, J.C. (1988) Estimating digestibility and oak browse diets for goats by in vitro techniques. Journal of Range Management, 41:255-258.
NEGI, S.S., PAL, R.N. and EHRICH, C. (1979) Tree Fodders in Himachal Pradesh. Eschborn, FRG: German Agency for Technical Cooperation.
NESER, J.A., COETZER, J.A.W., BOOMKER, J. and CABLE, H. (1982) Oak (Quercus rubor) poisoning in cattle. Journal of the South African Veterinary Association, 38(3):151-155.
NUNEZ-HERNANDEZ, G., HOLECHEK, J.L., WALLACE, J.D., GALYEAN, M.L., TEMBO, A., VALDEZ, R. and CARDENAS, M. (1989) Influence of native shrubs on nutritional status of goats: nitrogen retention. Journal of Range Management, 42(3):228-232.
OSTROWSKI, S.R., SMITH, B.P., SPIER, S.J., NORMAN, B.B. and OLIVER, M.N. (1989) Compensatory weight gain in steers recovered from oak bud toxicosis. Journal of the American Veterinary Medical Association, 195(4):481-484.
PEREVOLOTSKY, A., BROSH, A., EHRLICH, O., GUTMAN, M., HENKIN, Z. and HOLZER, Z. (1993) Nutritional value of common oak (Quercus calliprinos) browse as fodder for goats: experimental results in ecological perspective. Small Ruminant Research 11:95-106
RODER, W. (1985) Fodder tree use in Bhutan. In: WHITE, L. D. and TIEDEMAN, J.A. (eds.) Proceedings, International Rangeland Resource Development Symposium, Salt Lake City, Utah. pp33-41.
el-SABBAN, F.F., LONG, T.A. and BAUMGARDT, B.R. (1971) Utilization of oak sawdust as a roughage substitute in beef cattle rations. Journal of Animal Science, 32(4):749-755.
SHI, Z.C. (1988) Identification of the phenolic substances in bovine urine associated with oak leaf poisoning. Research in Veter�nary Science, 45:152-155.
SMITH, J. (1882) A Dictionary of Popular Names of Plants which Furnish the Natural and Acquired Wants of Man in all Matters of Domestic and General Economy. London: MacMillan and Co.
SPIER, S.J., SMITH, B.P., SEAWRIGHT, A.A., NORMAN, B.B., OSTROWSKI, S.R. and OLIVER, M.N. (1987) Oak toxicosis in cattle in Northern California: clinical and pathological findings. Journal of the American Veterinary Medical Association, 191(8):958-964.
UPHOF, J.C.T. (1968) Dictionary of Economic Plants. New York: Verlay von J. Cramer.
WILLIS, J.C. (1973) A Dictionary of the Flowering Plants and Ferns. 8th. edn. revised by AIRY-SHAW, H.K. London: Cambridge University Press.
Other titles in the Use of Trees by Livestock series include:
1 Prosopis
2 Acacia
3 Gliricidia
4 Anti-nutritive
factors
6 Cassia
7
Ficus
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Use of Trees by Livestock (NRI, 1994, 160 p.)
Use of Trees by Livestock: Ficus
R.T. Paterson and N.J.L. Clinch
Acknowledgements
© Crown copyright 1994
The Natural Resources Institute (NRI) is an internationally recognized centre of expertise on the natural resources sector in developing countries. It forms an integral part of the British Government's overseas aid programme. Its principal aim is to alleviate poverty and hardship in developing countries by increasing the productivity of their renewable natural resources. NRI's main fields of expertise are resource assessment and farming systems, integrated pest management, food science and crop utilization.
NRI carries out research and surveys; develops pilot-scale plant, machinery and processes; identifies, prepares, manages and executes projects; provides advice and training; and publishes scientific and development material.
Short extracts of material from this publication may be reproduced in any non-advertising, nonprofit-making context provided that the source is acknowledged as follows:
Paterson, R.T. and Clinch, N.J.L. (1994) Use of Trees by Livestock 7: Ficus. Chatham, UK: Natural Resources Institute.
Permission for commercial reproduction should be sought from:
The Head, Publishing and Publicity Services, Natural Resources Institute, Central Avenue, Chatham Maritime, Kent, ME4 4TB, United Kingdom.
No charge is made for single copies of this publication sent to the governmental and educational establishments, research institutions and non-profitmaking organizations working in countries eligible for British Government Aid. Free copies cannot normally be addressed to individuals by name but only under their official titles. Please quote LG7 when ordering.
Natural Resources Institute ISBN: 0 859544 367-6 Price £2.00
This booklet is printed on paper produced using the residues from sugar-cane processing.
Foreword
The importance of trees and shrubs in the feeding of animals in the tropics and sub-tropics has long been recognized by livestock owners. In and areas where the growth of herbaceous plants is limited by lack of moisture, leaves and edible twigs of trees and shrubs can constitute well over 50% of the biomass production of rangeland. At high altitudes, tree foliage may provide over 50% of the feed available to ruminants in the dry season, branches being harvested and carried to the animals. Even in regions of higher rainfall where grass supplies the major proportion of the dry matter eaten by ruminants, tree leaves and fruits can form an important constituent of the diet, particularly for small ruminants.
In the last two decades interest in the planting of trees as a source of feed for livestock has been encouraged by workers in research and development, but in contrast to the hundreds of indigenous species which are used as fodder, attention has focussed on a limited number of introduced species. Thus there are many publications reporting the chemical composition of Leucaena leucocephala leaves and suggesting management strategies for utilization of the tree for fodder, but it is more difficult to find information on alternative genera which might be equally, or more, appropriate.
The aim of this series of publications is to bring together published information on selected genera of trees which have the potential to increase the supply of fodder for ruminants. Each booklet summarizes published information on the fodder characteristics and nutritive value of one genus, with recommendations on management strategies, where available. Further, since the leaves of woody species frequently contain secondary compounds which may have an anti nutritional, or toxic, effect, a separate booklet summarizes the effects of a number of these compounds. It is hoped that the booklets will provide useful resource material for students, research and extension workers, interested in promoting the use of trees as a source of fodder for ruminants
Further copies of this booklet or others in the series can be obtained by writing to Publishing and Publicity Services at the Natural Resources Institute.
Margaret Gill
Livestock Production
Programme
Genus Ficus
Family MORACEAE
Principal species
Ficus auriculata (syn. F. roxburghii)
Ficus benghalensis
(syn. F. indica)
Ficus capens�s
FICUS carica
Ficus cunia
Ficus
elastica
Ficus elasticoides
Ficus exersperata
Ficus glaberrima
Ficus
glomerata
Ficus glumosa
Ficus gnaphalocarpa
Ficus hispida
Ficus
hookerii
Ficus iteophylla
Ficus lacor (syn. F. infectoria)
Ficus
ligens
Ficus nemoralis
Ficus neriifolia
Ficus pseudosycomorus
Ficus
religiosa
Ficus salicifolia
Ficus semicordata
Ficus sycomorus
Ficus
teleukat
Ficus thonningii
Ficus tsiela
Common names Fig
Earth
fig
Banyan
Peepul
Summary
Ficus is a large genus which is distributed widely throughout the warmer parts of Asia, Africa, America and Australia. It was introduced into the Mediterranean region thousands of years ago for the production of figs, which are consumed fresh, dried or preserved and used in cooking and for the production of wines and spirits. Apart from food for humans, members of the genus are used for many purposes. These include the production of fodder, latex, bird lime and wax, medicines for both humans and animals, fibres for ropes and cloth, wood for internal construction, packing, small tools and fuel, and shade for crops and settlements. Some species have religious significance in Africa and the Indian sub-continent. Many wild animals, ranging from insects to bats and monkeys, feed avidly on the fruits of a large number of species of figs, and so are responsible for the natural dissemination of the seed.
From the point of view of livestock production, many species are important for the provision of browse and fodder for cattle and buffaloes, small ruminants and camelids. In general terms, the foliage contains high levels of total digestible nutrients and is rich in calcium, reasonably high levels of crude protein and moderate amounts of crude fibre. The major nutritional problems lie in the low concentrations of phosphorus and the relative indigestibility of the protein fraction. While the fruits and foliage of most species are well accepted by livestock, F. elastica and F. tsiela are known to be toxic, particularly to cattle and buffaloes, where ingestion of large amounts of foliage can lead to animal mortality. The toxic principles have not been specifically determined, but proteolytic enzymes, coumarins, furo-coumarins and alkaloids are all known to occur in the genus. Some fig species are deciduous, but others retain their leaves all year round.
Seeds of Ficus spp. show low levels of germination, but vegetative propagation, either by stakes, including apicormic cuttings, or by tissue culture, is both rapid and efficient. Many species are attracting attention, particularly in India and Nepal, as multi-purpose trees in reafforestation projects.
Description and distribution
Ficus is a large and varied genus and comprises some 800 species. The following general information is taken mainly from Uphof (1968), Willis (1973) and Mabberley (1987).
While most commonly cultivated for fruit production in warm temperate and Mediterranean regions, Ficus is frequently found in areas with tropical and subtropical climates. Members of the genus range in growth habit from shrubs to large, spreading trees, although many species are rootclimbers, twiners and epiphytes. Adventitious and buttress roots are common and even the epiphytic members usually maintain contact with the soil through long aerial roots. Species such as F. benjamina climb up other trees, producing clasping roots which form a network that can eventually smother and kill the supporting tree. The banyan tree of India and Pakistan (F. benghalensis) is a crownspreading species which starts life as an epiphyte. Aerial roots reach the ground and become accessory trunks. In this way, a single individual may occupy an area of several hectares.
In southern Africa, Coates-Palgrave (1983) recognized three groups of figs as follows:
· strangler figs, the seeds of which will germinate in the forks of branches. The aerial roots overwhelm and eventually smother and kill the support tree;
· rock-splitting figs, which tend to grow on rocky outcrops. The roots squeeze into crevices and cracks, splitting apart even great boulders; and
· free-standing trees and shrubs. Strangler and rock splitting species can often grow as normal trees if the seeds germinate in open land.
Some members of the genus adopt a deciduous life-style. Thus, they are able to avoid unfavourable growing conditions by shedding their leaves and entering into a period of dormancy. While this has little influence on fruit yield, it can present special management problems when reliance is placed on the plants for animal feed. In addition to the direct loss of fodder, protein and digestible energy contents decrease and the fibre content increases in the period prior to the shedding of the leaves (Gohl, 1981).
The inflorescence of Ficus is hollow, consisting of a number of flowers (from 2 or 3, up to several thousand in the largest figs) inside a globular, or pear-shaped common recepticle, the narrow apical orifice of which is closed by small, overlapping bracts. Three types of flowers are produced: males, females, and sterile females known as gall-flowers. The sub-genera Urostigma, Sycomorus and Pharmacosycea are monoecious, all three types of flowers being produced on the same plant. In the dioecious sub-genus Ficus, male and gall-flowers are produced on one plant (gall-plant) and female flowers on another (seed-plant). In this case, both gall and seed plants must occur in the same locality for the formation of mature fruits. Pollination is carried out by insects which enter inside the fig to reproduce. For each species of Ficus, there would appear to be an individual gall-insect. These are often wasps belonging to one of several different genera of the hymenopterous family Agaonidae (Chalcidoidea). Many fig species bear their flowers in stalked bunches growing from old parts of the stem, although others fruit on underground stolons growing from the base of the trunk. These may be up to 10 m long, the figs being buried up to 10 cm deep in the soil. The seeds, carried within the fig, take the form of very small nuts which are naturally disseminated by the animals and birds which avidly eat the figs.
Many species of Ficus produce a milky latex which is sometimes tapped as a source of bird lime, or rubber.
Ficus may have originated in western Asia, the figs of commerce probably being introduced into the Mediterranean region thousands of years ago. While chiefly found in Indo-Malaysia and Polynesia, the genus is well represented in the warmer parts of Asia, western and southern Africa, Australia and the Americas from Mexico, Central America and the Caribbean into Peru and Brazil.
Fodder characteristics
In an early report from West Africa, Dalziel (1937) saw little economic merit in the genus Ficus, and of 35 species which he described, only F. iteophylla in N. Nigeria was considered to be of value as a fodder tree. More recently however, in Oyo State, Nigeria (humid forest and derived savannah zone), Carew et al. (1980) found F. exersperata compared very well with other indigenous browse species in terms of acceptibility. In northern Africa, in a Mediterranean climatic zone where browse is a more important component of rangeland than in the more tropical parts of the continent, Le Houerou (1980a) noted that F. carica, F. Iigens, F. pseudosycomorus, F. salicifolia and F. teleukat were palatable to sheep, goats and camels. In more eastern areas of Africa, leaves and fruits of F. thonningii, F. capensis and, to a lesser extent, F. glumosa, are rated as highly palatable to domestic animals (Audru, 1980; Le Houerou and Corra, 1980). Bark, leaves and twigs of F. sycomorus are consumed by elephants (Lamprey et al., 1980). In the Sudanian and Guinean zones, leaves of F. gnaphalocarpa are eaten by domestic livestock (Le Houerou, 1980b).

Table 1 - Proximate and fibre
analyses of Ficus spp.
In Nigeria, F. elasticoides is a browse species which is available at all times of the year. It can be fed fresh but the leaves have good milling characteristics and could be included in prepared rations if necessary (Asiegbu and Anugwa, 1988). Anugwa and Okori (1987) estimated the nutritive value of the foliage of the tree species F. elasticoides, Elaeis guineensis (oil palm) and Anacardium occidentale (cashew). They found that the fresh Ficus leaves and petioles were well accepted by West African dwarf lambs and led to higher levels of apparent digestibility than the other tree species, ranging from 70.1% for crude fibre (CF) and ether extract (EE) to 81.8% for nitrogen-free extract (NFE). Although the crude protein (CP) levels were modest, the apparent digestibility was high at 73.9%. It was suggested that feeding fig fodder to lambs should be actively encouraged in Nigeria.
Reference is frequently made to the use of Ficus in Asia as a dry season forage where branches are lopped and the leaves fed to both large and small ruminants (Gatenby et al., 1989a; Amatya, 1992; Upadhyay, 1992). In this region, several species of Ficus are highly rated as cattle feed. Gohl (1981) listed F. benghalensis, F. carica, F. glomerata, F. Iacor and F. religiosa as being trees which are often lopped for fodder. In an area of Nepal where the average mature fodder tree can be expected to produce 50-90 kg of fresh fodder per year (Panday, 1982), F. Iacor is capable of yielding some 150 kg of leaves and edible twigs. In a preliminary study with immature (4-5 year old) trees of F. glaberrima and F. semicordata, Amatya (1992) measured dry season yields of green matter of some 14 and 30 kg/tree respectively for the two species, of which 70% (November) to 80% (March) was leaf and the rest twigs.
In the hills of Nepal, numerous Ficus spp. show potential for bridging tile gap between the amount of feed needed by existing livestock populations and the availability from present feed resources (Gatenby et al., 1989a). In a listing of the flowering plants to be found in the country, Hara et al. (1982) noted the presence of 35 individual species of Ficus. Research work is being conducted on a number of these, including: F. semicordata, F. nemoralis, F. Iacor (syn. F. infectoria), F. cunia, F. auriculata (syn. F. roxburgii), F. glaberrima and others (Oli, 1987; Purohit, 1989; Dixit 1990). At the level of accepted usage, Gatenby et al. (1989b) listed F. roxburghii (syn. F. auriculata), F. Iacor, F. nemoralis and F. semicordata as important feed resources for buffaloes in the Koshi Hills.
It is interesting to note that in the Salija area of Nepal at an altitude of 1800-2300 m, where tree fodders are of great importance in the feeding of livestock, 86% of the women, but only 27% of the men interviewed in a study (Gold and Rusten, 1993) listed F. nemoralis as their first choice for tree fodder for animal feed. This species is frequently found on privately owned lands where it grows close to the houses and is thus readily accessible. It is a small, easily lopped tree without protective thorns. The favourite tree of the men in the area was Quercus semecarpifolia, a large, thorned species commonly found in the uncultivated areas further from the houses at higher altitudes. Although more dangerous and difficult to harvest, it is likely that a given quantity of fodder may be lopped more rapidly from the larger trees.
In a wider ranging survey of the Dhading and Bara districts of Nepal (Upadhyay, 1991; Kapali, 1992), F. neriifolia was ranked second behind Castanopsis tribuloides as the most valued tree for animal production. No preferences relating to the gender of the livestock owners were recorded in this study.
A number of reports refer to the nutritive value of Ficus spp. (Mia et al., 1960; Gohl, 1981; Chanda and Bhaid, 1987) and representative data are shown in Table 1. There is considerable variation both between and within species but this is due, at least in part, to the effect of season and to the deciduous growth pattern of many species within the genus. Further variation may be explained by differences in genetic potential, bioclimatic conditions and cropping systems. There would also appear to be differences between species of domestic livestock in the way in which they can utilize fig foliage. Goats utilized the nitrogen content of F. religiosa more efficiently than cattle (Mia et al., 1960) and they were able to survive on a diet of F. tsiela, a species which is reportedly toxic to cattle and buffaloes (Divakaran Nair et al., 1985). In common with many tree fodders, those species of Ficus which do not shed their leaves tend to maintain their quality well into the dry season, making them valuable feed resources when herbaceous vegetation is in short supply (Nehra et al., 1987; Gatenby et al., 1989a,b).
Goats appear to consume fresh fodder of Ficus spp. in relatively larger quantities than do cattle. To compare intake between animals of widely differing sizes, the amount of feed consumed may be expressed on the basis of daily voluntary intake of fodder dry matter per 100 kg of animal liveweight. Using this measure, goats were reported to consume 3.2 kg of F. hookerii (Gupta and Balaraman, 1989), while in a single, short-term experiment with F. religiosa, goats ate 5.2 kg, compared with 2.0 kg by cattle (Mia et m., 1960). It is, however, uncommon for animals to be offered tree fodder alone unless no other feed sources are available. In the arid Haryana region of India, recommended daily feeding levels of fresh F. religiosa leaves and edible twigs are 1-2 kg/head for goats, 1-1.5 kg for sheep, and 3-4 kg for camels. The order of decreasing acceptance of tree fodders in general is considered to be goats, sheep and camels, with cattle showing least preference for browse (Nehra et al., 1987).
Rusten and Gold (1991), reviewing indigenous knowledge regarding tree fodder in Nepal, noted a number of species, including F. nemoralis, which could increase both milk production and butterfat content when fed to lactating cattle. Other species such as F. hispida, F. auriculata and F. Iacor are considered by many authors to be useful dietary components for both large and small ruminants. There are, however, few data available concerning the long-term effects of feeding fodder of Ficus spp. to livestock. Most of the work reported in the literature has been short-term, aimed at the study of metabolic balances over a feeding period of one or two weeks. The growth rates measured in such work do not necessarily bear a close relationship to animal productivity over a full season.
In an experiment with mature draught oxen in the Koshi Hills region of Nepal, animals with free access to rice straw and 4 kg/day of freshly harvested leaves of F. auriculata, consumed more feed and regained more weight in the three weeks following a two-week period of work, than animals fed on rice straw alone. The tree fodder did not prevent the loss of some 8 kg/head during the work period but it allowed the oxen to recover the lost body condition in the following two to three weeks. Although both rice straw and tree foliage were considered to be deficient in nitrogen available to the rumen microbes, the author (Pearson, 1990) speculated that the tree fodder could be an acceptable alternative to expensive concentrates for working oxen kept on a diet based on poor quality roughage.
Murugan and Kathaperumal (1987) compared different supplements to sheep fed on a standard, low quality basal diet. Those animals with access to F. religiosa leaves gained 4.07 kg in weight over a post-weaning period of 84 days (48 g/day) while those supplemented with Azadirachta indica and Enterolobium saman leaves lost 0.53 kg and 0.23 kg respectively in the same period.
In the Nigerian work of Anugwa and Okori (1987) mentioned above, West African dwarf lambs gained 71 g/day over a 14-day period when fed a sole diet of F. elasticoides foliage. In a more recent study using the same fodder species and breed of sheep (Anugwa, 1990), the high nutritive value shown by the levels of digestible energy (12.26 MJ/kg DM), total digestible nutrients (68.8%) and digestible protein (11.5%), indicated the potential of the fodder for dry season ruminant production.
In the Mhow area of India, where it is common practice to feed goats on the leaves of trees gathered in the forest, an experiment was conducted with uncastrated male goats aged about 14 months and weighing some 25 kg. A diet of dried leaves of F. tsiela was fed together with 0.3 kg/day of wheat bran and 5 g of common salt. The animals consumed the tree fodder dry matter at an acceptable daily rate of 2.52 kg/100 kg body weight, and gained a modest average of 42 g/day liveweight. The leaves were relatively high in calcium (3.43%) but low in phosphorus (0.09%). The digestibility of the dry matter was estimated at 50.1% but the crude protein was almost totally indigestible and the feed was classified as inferior quality roughage (Chanda and Bhaid, 1987).
In a further study in the same area, F. tsiela was shown to be less economical for the feeding of goats than other tree species such as Hardwickia binata, which could be fed without wheat bran supplements. Bundles of tree fodder offered for sale in the Patti Bazar included Ficus samples which consisted of 55% leaf and 45% twig and H. binata samples which consist ed of 75% leaf and only 25% twig. Since the price of the bundle did not vary with the fodder species, the daily feeding cost using Ficus and wheat bran was calculated at 163% higher than with the H. binata alone (Mehta and Bhaid, 1990).
In the Sikkim area of India, where trees commonly grown for fodder include a number of Ficus spp. (Bernet, 1985), dry matter of F. hookerii was consumed by local goats (average weight 25.6 kg) at a level of 3.2 kg/100 kg liveweight. The digestibility of the dry matter and crude protein were estimated at 54.3 and 35.7% respectively. The high level of calcium (3.38%) and the lower level of phosphorus (0.21%) were not dissimilar to those noted above for F. tsiela and animals showed positive balances for nitrogen and calcium, and a marginal negative balance for phosphorus when fed on a sole diet of the fig leaves. It was noted that F. hookerii, when supplemented with bone meal or minerals, could provide a maintenance diet for goats, although in view of the presence of anti-nutritive factors (possibly tannins), long-term feeding trials should be conducted to study potentially adverse effects (Gupta and Balaraman, 1989).
Although conclusive, long-term data are scarce, the evidence suggests that while the digestibility of the protein component, and the level of phosphorus in Ficus spp. may both be relatively low, the protein and calcium contents are high enough, at least in those species which are commonly utilized by livestock, to make them valuable as dry season forage.
Anti-nutritive factors
Although Panday (1982) has implicated F. auriculata in decreases in milk production, dysphagia and urinal disorders in bovines, only F. tsiela and F. elastica are commonly reported as causing specific toxic effects in domestic livestock.
Gohl (1981) noted that while several species of Ficus were commonly lopped for animal feed, particularly in India and Pakistan, the young leaves of F. elastica were poisonous to most animals, causing profuse sweating, paralysis and eventual death.
Mortality in cattle and buffaloes due to consumption of the leaves of F. tsiela has been reported from a number of areas of India (Rajan et al., 1986), although as noted above, goats are able to grow, at least for a limited period, on a diet of foliage from this species when it is supplemented with 0.3 kg/day of wheat bran (Chanda and Bhaid, 1987). The foliage, which is sometimes stored on farms for use as a manure for paddy fields (Divakaran Nair et al., 1985), is readily eaten by livestock and toxicity has been provoked in young calves (2-3 months of age) by the consumption of a total of 2.5 kg of mature, tender leaves and soft stems over a period of three days, or by 3-4 kg spread over some 12 days. Physical symptoms, which occur within 4-12 days after ingestion of the fodder, include excessive salivation, frequent bellowing, laboured breathing and intermittent tonic convulsions. The symptoms are predominantly nervous in nature and associated with significant histological changes, including cerebral oedema and demyelination, together with renal and hepatic degeneration and necrosis (Divakaran Nair et al., 1985; Rajan et al., 1986). Pathological studies suggest that the leaves contain substances which predominantly affect the capillaries of the central nervous system.
Several potentially toxic substances have been isolated from Ficus spp. The proteolytic enzyme ficin is found in the latex of several members of the genus, including F. carica, the common fig. This compound is more toxic when administered intravenously than when ingested, and the toxicity depends primarily on the amount given, rather than on the concentration of the dose. Sub-lethal doses provoke vomiting, sanguineous diarrhoea and general prostration.
A furo-coumarin, ficusin (also known as psoralene), has been isolated from the leaves of F. carica, and a dialysable, alkaloid-like compound has been found in the latex. This species is known to cause percutaneous photosensitization in man, probably due to the presence of coumarins and furocoumarins, including ficusin (Watt and Breyer Brandwijk, 1962).
The leaves of F. septica, which are used in Papua New Guinea to cure fevers, fungal and bacterial diseases, contain a range of alkaloids, including tylophorine, septicine and autofine. A recent isolate is the new indolizidine alkaloid, ficuseptine.
Methanolic extracts of the leaves displayed intense antibacterial and antifungal activities (Baumgartner et al., 1990). While many alkaloids are beneficial to both humans and livestock due to their medicinal properties, others are known to be toxic, the effect varying with the species of animal (Kingsbury, 1964; Southon and Buckingham, 1989).
Although precise identification of the toxic principles has yet to be made, it is likely that one or more of the chemical groups noted above could be involved in the reported animal mortality resulting from ingestion of F. elastica and F. tsiela.
It has been noted above that the protein content of the foliage of Ficus spp. is often poorly digested by livestock. Some fig species, such as F. glumosa, have been used for the tanning leather (Uphof, 1968), and it is well known that tannin, the active fraction in the process, is capable of interfering with the digestion of proteins (Mangan, 1988). Although published confirmation is lacking, it would seem likely that tannins are responsible for the occasional poor utilization of fig fodder.
The leaves of F. exersperata contain a high proportion of calcium silicate (Watt and BreyerBrandwijk, 1962), an abrasive which may be capable of causing intestinal inflamation and associated health problems in both cattle and humans.
Management
In common with many plants that produce tiny seeds, the germination of Ficus spp. is often poor and F. Iacor, an important fodder tree in Nepal at altitudes between 800 and 2000 m, has a germination rate of only 15% (Jackson, 1987). While this is adequate for natural regeneration, it presents problems for reafforestation activities. In such a programme in the central Himalayas of India, eight species of Ficus have been chosen for distribution to villagers as multi-purpose trees, by virtue of their resistance to drought and frost, high photosynthetic rates and efficiency of water usage. Fortunately, vegetative propagation is easy and both seedlings and stem cuttings are produced by the project for planting on small farms for use as shade, fodder, fuel wood and edible figs for jams and curries (Purohit, 1989).
Several techniques of vegetative propagation have been successfully utilized, particularly in the Indian sub-continent. In the middle hills region of Nepal, where F. nemoralis is highly palatable to grazing animals, stem cuttings of about 1 m length are planted within the canopy of Neolitsea umbrosa, a small bushy tree that is totally unattractive to livestock and therefore ignored by them. The fig is protected from browsing as it grows up through the crown of the nurse tree (Gold and Rusten, 1993). Although no data are presented, farmers claim that the fig grows more quickly when associated with N. umbrosa, a species which is used locally for fuel wood, tools and utensils, and whose fruit is reputed to have medicinal properties.
In Senegal and Madagascar, a number of Ficus spp. including F. thonningii have been successfully propagated using apicormic shoots. These are large shoots which grow vertically upward from lateral branches and which exhibit many of the growth characteristics of mature seedlings. When cut at an age of about three years, they have a basal diameter of some 15 cm and are then trimmed to a length of 2.5 m. After cutting, they are laid horizontally in the shade for one week and then stood vertically for a further three weeks to harden before being planted to a depth of about 50 cm. These large cuttings establish rapidly and require little protection from browsing. The technique is well suited to the planting of trees as living fence posts, or in existing pastures (Jolin and Torquebiau, 1992).
An in vitro micropropagation technique was developed in Nepal for F. Iacor, permitting multiplication by a factor of 20-30 every 8-12 weeks over a period of at least two years (Amatya and Rajbhandary, 1991). Shoot tip explants from standing trees were treated with benzyl aminopurine, kinetin and casein hydrolysate to promote shoot proliferation. Microshoots were rooted in sand at high humidity under a polythene cover. When rooted plants 5-6 cm high were transferred to soil for hardening prior to planting out in the field, no losses were suffered. Micropropagation offers the potential for cheap, rapid multiplication of disease-free planting stock from elite, individually selected parent trees.
In much of the area where Ficus spp. are important for the feeding of livestock, individual holdings are small and are not capable of supporting all of the nutritional needs of the animals kept on them. In India and Nepal, it is common practice to care for trees planted along the boundaries, or within the confines of the family property, but trees growing on common land and in communal forests are also harvested, either by the owners of the livestock or by others, in order to sell fodder to stockmen (Mehta and Bhaid, 1990). The newly opened leaves of F. nemoralis, one of the most popular fodder species in the Salija area of Nepal, are nutritious, but are known to produce watery dung. Mature leaves, on the other hand, lead to more solid manure which is easier to collect and handle. Mature foliage is therefore harvested selectively where the manure from livestock is valued as an important resource for use within the cropping activities of the farm (Rusten and Gold, 1991).
In the Haryana region of India, with light soils and an annual rainfall of 250-500 mm most of which falls in about three months, naturally occurring F. religiosa is an important fodder tree. Trees on private land are usually protected but those growing along roadsides and on common grazing lands are often subject to over-use. Stockmen harvest foliage from all trees which are known to be palatable to livestock. The cut material may be fed to herded animals in situ, or carried back to stock kept in confinement. In times of plenty, surplus feed is cut for feeding later in the year. Under these arid conditions, lopping once per year of a single tree of F. religiosa will yield 150-300 kg of fresh, edible fodder, compared with 15-50 kg/tree for other local fodder species, including members of the genera Acacia, Prosopis and Zizyphus (Nehra et al., 1987). Optimum management practices and the resultant productivity of a particular tree will depend upon those variables which govern speed of plant growth such as soil fertility, rainfall, temperature, competition and biotic factors. In the arid Haryana region, however, where cutting should be minimized in areas prone to soil erosion, the following lopping recommendations have been made to ensure both the survival and the sustainable productivity of the existing tree fodder resource (Nehra et al., 1987):
· only trees over 2 m tall, and branches over 7.5 cm in diameter should be lopped;
· only the lower two-thirds of the foliage should be removed;
· only mature foliage should be cut in a rotational lopping system, such that each branch is only cut once per year.
Alternative uses
As multi-purpose trees, members of the genus Ficus are used in a variety of ways in both subsistence and commercial agriculture (Watt and Breyer-Brandwijk, 1962; Uphof, 1968; Willis, 1973; Mabberly, 1987).
Perhaps the most important use is in the production of fruits which may be eaten raw, dried or preserved, used in cooking or for the preparation of wines and spirits. F. carica, the best known commercial fig, is probably native to southwest Asia but it has been cultivated in the Mediterranean region for some 6000 years. Many named varieties have been developed in a number of countries, and have been classified either according to the colour of the fruit, which may be deep purple or greenish yellow, or with the principal end use, such as fresh, dried, or preserved fruit. In Africa, America and Asia, many species other than F. carica produce fruits which are gathered for human consumption. These include F. auriculata, F. capensis, F. cunia, F. padifolia, F. palmata, F. pumila and F. sycomorus. At high altitudes in India, fruit yields of 112 kg/tree/year were reported from F. recemosa (Purohit, 1989).
Although the importance of latex production has decreased in recent decades due to the cultivation of the South American species Hevea brasiliensis and the development of synthetic compounds, a number of Ficus spp. have been tapped for rubber production. These include F. elastica (Assam, or India rubber), F. vogelii (Dahomey rubber), F. nekbuda (in the Congo) and F. annulata (in Australia and Malacca). Allied to the production of sticky latex, a number of species such as F. anamani, F. benghalensis and F. glumosa yield bird lime, which is used to trap small birds.
The bark of many species such as F. artocarpoides, F. glumosa, F. Iingua, F. Iuteola, F. michelsonii and F. vogelii is used to make cloth, while the bark and twigs of F. baron) and F. tinctoria produce fibres used for ropes and fishing nets.
In Java, wax is extracted from F. ceriflua and F. tox�caria. This is used in batik work and also for the making of candles. In southeast Polynesia, a red dye is extracted from the fruit of F. tinctoria.
In several parts of the world, Ficus spp. are used in traditional medicines. The latex and bark of F. anthelmintica are used to treat fevers in Peru, while the same plant parts of F. cotinifolia are used to treat wounds and bruises in Mexico and Central America. The latex of F. benghalensis is used for bruises, rheumatism and lumbago in Burma and Malaysia. A decoction of leaves of F. crassinervia finds a use in the treatment of liver diseases in Cuba. F. septica acts as an antidote to poisoning resulting from eating or touching some toxic species of fish and crabs in Malaysia. F. involuta and F. Iacor are used in a range of medicines in Mexico and China respectively, while in India F. hispida is used in the treatment of jaundice, leprosy and anaemia (Acharya and Kumer, 1984). The latex of a number of species, including F. carica, shows anthelmintic properties associated with the presence of the proteolytic enzyme ficin.
The rough leaves of F. exersperata can be employed as a sandpaper to polish wood, while the bark of F. glumosa is used in the tanning of leather in parts of Africa.
The wood of Ficus spp. is often whitish grey in colour. It is light and fairly hard, and not usually durable, although timber from some species withstands prolonged immersion in water. It is of limited commercial value, used principally for cheap furniture, interior construction, tea chests, yokes and carvings. Species such as F. benghalensis, F. glomerata, F. Iaevigata and F. mucoso are employed in this way. Planks are cut from the buttress roots of F. sycomorus.
Wood from lopped branches is often valued as a source of fuel.
Silkworms are fed on the leaves of Ficus spp. In India, Theophila religiosue is raised on leaves of F. religiosa and F. benghalensis, while in the northwest Himalayas, Ocinara signifera is grown on leaves of F. tsjuleka. Lac insects, kept for the production of shellac, are grown on F. religiosa.
Both F. religiosa (peepul) and F. benghalensis (banyan) have religious significance for Hindus and Buddhists, while in East Africa, F. capensis is revered by the Kikuyu people as sacred shrines or places of sacrifice to ancestral spirits.
Several species such as F. macrophylla (Moreton Bay fig) and F. thonningii are large, attractive, spreading trees which are often planted for shade in avenues and gardens in Australia and tropical Africa respectively. Other species are used to provide shade for crops and domestic animals and mark the boundaries of plots in the Indian sub-continent and Southeast Asia (Audru, 1980).
References and further reading
ACHARYA, B.M. and KUMER, K.A. (1984) Chemical examination of the bark of Ficus hispida Linn. Current Science, India, 53:1034-1035.
AMATYA, N. and RAJBHANDARY, S.B. (1991) In vitro propagation of Ficus lacor Buch.-Ham. Banko Janakari, 3(1):17-20.
AMATYA, S.M. (1992) Foliage biomass yield from three species of fodder trees in Nepal: a preliminary study. Banko Janakari, 3(3):19-20.
ANUGWA, F.O.I. (1990) Feed intake, nutrient digestibility and nitrogen balance by sheep fed leaves of Panicum maximum, Calopogonium mucunoides and Ficus elasticoides. Bulletin of Animal Health and Production in Africa, 38:113- 118.
ANUGWA, F.O.I. and OKORI, A.U. (1987) The nutritive value of three Nigerian browse plants eaten by sheep. Bulletin of Animal Health and Production in Africa, 35:223-228.
ASIEGBU, J.E. and ANUGWA, F.O.I. (1988) Seasonal availability, physical and chemical characteristics of four major browse plants used for stall-feeding of livestock in eastern Nigeria. Bulletin of Animal Health and Production in Africa, 36:221-228.
AUDRU, J. (1980) Ligneous and subligneous forage and fruit trees in the Guinean zone: prospects for utilization in animal production. In: LE HOUEROU, H.N. (ed.) Browse in Africa: the Current State of Knowledge. Addis Ababa, Ethiopia: International Livestock Centre for Africa (ILCA).
BAUMGARTNER, B., ERDELMEIER, C.A.J., WRIGHT, A.D., RALI, T. and STICHER, O. (1990) An antimicrobial alkaloid from Ficus septica. Phytochemistry, 29:3327-3330.
BENNET, S.S.R. (1985) Ethnobotanical studies in West Sikkim. Journal of Economic and Taxonomic Botany, 7:317-321.
CAREW, B.A.R., MOSI, A.K., MBA, A.U. and EGBUNIKE, G.N. (1980) The potential of browse plants in the nutrition of small ruminants in the humid forest and derived savanna zones of Nigeria. In: LE HOUEROU, H.N. (ed.) Browse in Africa: The Current State of Knowledge. Addis Ababa, Ethiopia: International Livestock Centre for Africa (ILCA).
CHANDA, S.K. and BHAID, M.U. (1987) Studies on the digestibility and nutritive value of common dried fodder tree leaves of M.P. on goats: 4. Hay of phephar tree leaves (Ficus tsiela Roxb). Livestock Adviser, 12(3):5-8.
COATES-PALGRAVE, K. (1983) Trees of Southern Africa. 2nd edn. Cape Town: C. Struik.
DALZIEL, J.M. (1937) The Useful Plants of West Tropical Africa. London: Crown Agents for the Colonies.
DIVAKARAN NAIR, N., VALSALA, K.V., RAMACHANDRAN, K.M. and RAJAN, A. (1985) Experimental studies on chela leaf (Ficus tsiela Roxb) poisoning in calves. Kerala Journal of Veterinary Science, 16:94-99.
DIXIT, P. (1990) Implementing farm-forestry practices in Nepal: experiences and achievements. Forestry Chronicle, 66:242-245.
GATENBY, R.M., MAHATO, S.N. and SHRESTHA, N.P. (1989a) Animal Production in the Hills of Nepal. PAC Technical Paper 112. Dhankuta, Nepal: Pakhribas Agricultural Centre (PAC).
GATENBY, R.M., NEOPANE, S.D. and CHEMJONG, P.B. (1989b) Traditional Feeding Practices for Buffaloes in the Koshi Hills. PAC Technical Paper 99. Dhankuta, Nepal: Pakhribas Agricultural Centre (PAC).
GOHL, B. (1981) Tropical Feeds: Feed Information Summaries and Nutritive Values. FAO Animal Production and Health Series No.12. Rome: Food and Agriculture Organization (FAO).
GOLD, M.A. and RUSTEN, E.P. (1993) Learning from the farmers of Nepal. Agroforestry Today, 5(1):10-12.
GUPTA, H.K. and BALARAMAN, N. (1989) Nutritive value of nevaro (Ficus hookerii) leaves for goats. Indian Journal of Animal Sciences, 59:186-187.
HARA, H., CHATER, A.O. and WILLIAMS, L.H.J. (1982) An Enumeration of the Flowering Plants of Nepal, vol. 3. London: British Museum (Natural History).
JACKSON, J.K. (1987) Manunl of Afforestation in Nepal. Kathmandu, Nepal: Nepal-UK Forestry Research Project.
JOLIN, J. and TORQUEBIAU, E. (1992) Large cuttings: a jump start for tree planting. Agroforestry Today, 4(4):15-16.
KAPALI, S.P. (1992) Statistical analysis of most used fodder tree species. Banko Janakari, 3(3):13-16.
KINGSBURY, J.M. (1964) Poisonous Plants of the United States and Canada. Englewood Cliffs, USA: Prentice Hall Inc.
LAMPREY, H.F., HERLOCKER, D.J. and FIELD, C.R. (1980) Report on the State of Knowledge on Browse in East Africa in 1980. In: LE HOUEROU, H.N. (ed.) Browse in Africa: the Current State of Knowledge. Addis Ababa, Ethiopia: International Livestock Centre for Africa (ILCA).
LE HOUEROU, H.N. (1980a) Browse in northern Africa. In: LE HOUEROU, H.N. (ed.) Browse in Africa: the Current State of Knowledge. Addis Ababa, Ethiopia: International Livestock Centre for Africa (ILCA).
LE HOUEROU, H.N. (1980b) The role of browse in the Sahelian and Sudanian zones. In: LE HOUEROU, H.N. (ed.) Browse in Africa: the Current State of Knowledge. Addis Ababa, Ethiopia: International Livestock Centre for Africa (ILCA).
LE HOUEROU, H.N. and CORRA, M. (1980) Some browse plants of Ethiopia. In: LE HOUEROU, H.N. (ed.) Browse in Africa: the Current State of Knowledge. Addis Ababa, Ethiopia: International Livestock Centre for Africa (ILCA).
MABBERLEY, D.J. (1987) The Plant Book. New York: Cambridge University Press.
MANGAN, J.L. (1988) Nutritional effects of tannins in animal feeds. Nutrition Research Reviews, 1:209-231.
MECHA, I. and ADEGBOLA, T.A. (1980) Chemical composition of some southern Nigerian forage eaten by goats. In: LE HOUEROU, H.N. (ed.) Browse �n Africn: the Current State of Knowledge. Addis Ababa, Ethiopia: International Livestock Centre for Africa (ILCA).
MEHTA, M.K. and BHAID, M.U. (1990) Economics of fodder tree leaves feeding to goats. Livestock Adviser, 15(12):38-39.
MIA, W.H., MAJUMDAR, B.N., SAHAI, B. and KEHAR, N.D. (1960) Studies on tree leaves as cattle fodder: 4. The nutritive value of Pipal leaves (Ficus religiosa). Indian Journal of Dairy Science, 13:9-15.
MURUGAN, M. and KATHAPERUMAL, V. (1987) Utilization of tree fodder in sheep. Indian Journal of Animal Sciences, 57:1145-1147.
NEHRA, O.P., OSWAL, M.C. and FARODA, A.S. (1987) Management of fodder trees in Haryana. Indian Farming, 37(3):31, 33.
OLI, K.P. (1987) Small Ruminant Production in the Easten' Hills of Nepal. PAC Working Paper 11. Dhankuta, Nepal: Pakhribas Agricultural Centre (PAC).
PANDAY, K.K. (1982) Fodder Trees and Tree Fodder in Nepal. Switzerland: Swiss Development Corporation, Berne/Swiss Federal Institute of Forestry Research, Birmensdorf.
PEARSON, R.A. (1990) A note on live weight and intake and digestibility of food by draught cattle after supplementation of rice straw with the fodder tree Ficus auriculata. Animal Production, 51:635-638.
PUROHIT, A.N. (1989) Indians testing Ficus as high altitude MPT. Agroforestry Today, 1(3):19-20.
RAJAN, A., DIVAKARAN NAIR, N., VALSALA, K.V., MARYAMMA, K.I. and RAMACHANDRAN, K.M. (1986) Pathology of a nervous disorder in cattle caused by the toxicity of the leaves of the tree Ficus tseila Roxb. ludian Veterinary Journal, 63:184-186.
ROSE-INNES, R. (1967) Land and Water Survey in the Upper and Northern Regions, Ghana. Final Report, vol IV. FAO/SF:31 GHA. Rome: Food and Agriculture Organization (FAO).
RUSTEN, E.P. and GOLD, M.A. (1991) Understanding an indigenous knowledge system for tree fodder via a multi-method on-farm research approach. Agroforestry Systems, 15:139-165.
SOUTHON, I.W. and BUCKINGHAM, J. (1989) Dictionary of Alkaloids. London: Chapman and Hall. 2 vols.
UPADHYAY, L.R. (1991) The Country-wide Survey of Farmers Knowledge and Perceptions about Tree Fodder1990 (The Pilot Surveys: Dhading and Bara Districts). FRD Occasional Paper No.1/91. Kathmandu, Nepal: Forest Research Division.
UPADHYAY, L.R. (1992) Use of fodder trees in Jhapa and Sunsari Districts in the eastern Terai. Banko Janakari, 3(3):17-18.
UPHOF, J.C. Th. (1968) Dictionary of Economic Plants. 2nd edn. New York: Verlay von J. Cramer.
WATT, J.M. and BREYER-BRANDWIJK, M.G. (1962) Medicinal and Poisonous Plants of Southern and Eastern Africa. 2nd edn. Edinburgh: Collins.
WILLIS, J.C. (1973) A Dictionary of the Flowering Plants and Ferns. 8th edn. Cambridge, UK: University Press.
Other titles in the Use of Trees by Livestock series include:
1 Prosopis
2 Acacia
3 Gliricidia
4 Anti-nutritive
factors
5 Quercus
6
Cassia
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Use of Trees by Livestock (NRI, 1994, 160 p.)
Use of Trees by Livestock: Calliandra
R.T. Paterson
Acknowledgements
© Crown copyright 1994
The Natural Resources Institute (NRI) is an internationally recognized centre of expertise on the natural resources sector in developing countries. It forms an integral part of the British Government's overseas aid programme. Its principal aim is to alleviate poverty and hardship in developing countries by increasing the productivity of their renewable natural resources. NRI's main fields of expertise are resource assessment and farming systems, integrated pest management, food science and crop utilization.
NRI carries out research and surveys; develops pilot-scale plant, machinery and processes; identifies, prepares, manages and executes projects; provides advice and training; and publishes scientific and development material.
Short extracts of material from this publication may be reproduced in any non-advertising, nonprofit-making context provided that the source is acknowledged as follows:
Paterson, R.T. (1994) Use of Trees by Livestock 8: Calliandra. Chatham, UK: Natural Resources Institute.
Permission for commercial reproduction should be sought from:
The Head, Publishing and Publicity Services, Natural Resources Institute, Central Avenue, Chatham Maritime, Kent, ME4 4TB, United Kingdom.
No charge is made for single copies of this publication sent to governmental and educational establishments, research institutions and nonprofit making organizations working in countries eligible for British Government Aid. Free copies cannot normally be addressed to individuals by name but only under their official titles. Please quote LG8 when ordering.
Natural Resources Institute
ISBN: 0 85954 369-2
Price
£2.00
This booklet is printed on paper produced using the residues from sugar-cane processing.
Foreword
The importance of trees and shrubs in the feeding of animals in the tropics and sub-tropics has long been recognized by livestock owners. In arid areas where the growth of herbaceous plants is limited by lack of moisture, leaves and edible twigs of trees and shrubs can constitute well over 50% of the biomass production of rangeland. At high altitudes, tree foliage may provide over 50% of the feed available to ruminants in the dry season, branches being harvested and carried to the animals. Even in regions of higher rainfall where grass supplies the major proportion of the dry matter eaten by ruminants, tree leaves and fruits can form an important constituent of the diet, particularly for small ruminants.
In the last two decades interest in the planting of trees as a source of feed for livestock has been encouraged by workers in research and development, but in contrast to the hundreds of indigenous species which are used as fodder, attention has focused on a limited number of introduced species. Thus there are many publications reporting the chemical composition of Leucaena leucocephala leaves and
suggesting management strategies for utilization of the tree for fodder, but it is more difficult to find information on alternative genera which might be equally, or more, appropriate.
The aim of this series of publications is to bring together published information on selected genera of trees which have the potential to increase the supply of fodder for ruminants. Each booklet summarizes published information on the fodder characteristics and nutritive value of one genus, with recommendations on management strategies, where available. Further, since the leaves of woody species frequently contain secondary compounds which may have an anti-nutritional, or toxic, effect, a separate booklet summarizes the effects of a number of these compounds. It is hoped that the booklets will provide useful resource material for students, research and extension workers, interested in promoting the use of trees as a source of fodder for ruminants.
Further copies of this booklet or others in the series can be obtained by writing to Publishing and Publicity Services at the Natural Resources Institute.
Margaret Gill Livestock Production Programme
Genus Calliandra
FamilyLEGUMINOSAE
SubfamilyMIMOSOIDEAE
TribeINGEAE
Principal species
Calliandra angustifolia
Calliandra calothyrsus (syn. C.
confusa)
Calliandra haematocephala (syn. C. inaequilatera)
Calliandra
houstoniana (syn. C. houstoni, C. houstonii)
Calliandra
pittieri
Calliandra portoricensis (syn. Zapoteca portoricensis)
Calliandra
surinamensis
Calliandra tetragolla (syn. Zapoteca tetragona)
Common names
Powder puff tree (or shrub)
Cabellos de angel (Central
America)
Lehua haole
(Hawaii)
Summary
Calliandra is a large genus of mainly tropical American shrubs and small trees. Many species produce attractive flowers and are valued by honey bees. Others are used for shade, green manure, reafforestation, fuelwood, charcoal and timber. Recently, attention has focused on its potential as a source of fodder for livestock, but published information is largely restricted to a single species, C. calothyrsus.
This species tolerates a wide range of soil conditions and persists in regions of low rainfall although it does best under 2000-4000 mm/year. It coppices readily and will tolerate frequent defoliation. It appears to be resistant to most pests and diseases and may become a useful alternative to Leucaena leucocephala in areas where psyllids are a problem.
The leaves of C. calothyrsus contain 20-22% crude protein and reasonable levels of minerals. It has no reported toxic effects on higher animals but its feeding quality is reduced by high levels of condensed tannins. Animal acceptance and digestibility are variable. It should be seen as a supplement for poorquality roughage, and should be managed under systems of direct browsing or daily cutting.
Description and distribution
Calliandra is a large genus in which there is currently a degree of taxonomic confusion and synonymy (NRC, 1983). It is variously reported to consist of over 100 (NRC, 1983), some 150 (Allen and Allen, 1981), or up to 200 (Mabberley, 1987) species. It has recently been suggested that the genus should be split and species such as C. portoricensis and C. tetragona be consigned to the genus Zapoteca. This view is now generally accepted but for the purposes of this booklet they are treated as remaining within Calliandra. Most published information refers to C. calothyrsus but even within this species there is such a high degree of variation in agronomic characteristics and soil requirements that the accepted taxon could be considered a complex of more than one species (Netera et al., 1992).
Members of the genus are frequently unarmed and usually exist as small trees or shrubs, although there are occasional large trees and perennial herbs. They are normally straggling, highly branched plants with bipinnate (compound) leaves which have a tendency to fold at night. The showy, attractive flowers, which are usually either red or white in colour, often resemble pompoms or powder puffs and give rise to the common name of the group-powder puff shrubs or trees. The pods are flat with raised margins, narrow at the base, and are elastically dehiscent from the apex. The seeds, up to 15 in a pod, are laterally compressed and germinate readily.
Calliandra spp. occur in a wide variety of habitats ranging from rainforest to dry mountain slopes. They are usually found at altitudes up to 1500-1800 m but some species such as C. grandiflora occur at up to 3000 m (Macqueen, 1993). Some species, and even accessions within species, appear to require fertile soils of neutral reaction, while others will tolerate infertile, acid soils with a high level of aluminium saturation. They are often good colonizers of denuded areas and will tolerate soils which are heavily compacted and poorly aerated. They persist in poorly drained, sloping, podzolic soils (Gutteridge, 1990), although Shelton et al. (1991) considered them to be moderately intolerant of waterlogging. They have a marked capacity to reduce soil erosion and have been successfully used for reafforestation in many areas (Allen and Allen 1981; NRC, 1983; Shelton et al., 1991).
Nodulation has been validated and nitrogen fixation is assumed in 11 Calliandra spp. (Allen and Allen, 1981; Brewbaker et al., 1990). Although estimates of the quantity of nitrogen fixed per year are largely unavailable, trees show little response to fertilizer nitrogen and they would therefore appear to be self-sufficient in nitrogen in most environments. Both fast and slow growing strains of Rhizobium have been isolated from Calliandra root nodules, and the genus appears to be promiscuous with regard to nodulation (NRC, 1983). There are reports of nonnodulating members of the genus but they are rarely substantiated and should therefore be treated with caution (Allen and Allen, 1981). The roots of naturally occurring strands of C. calothyrsus are usually infected by beneficial mycorrhizal fungi which assist in the absorption of phosphorus and other nutrients (NRC, 1983; Shelton et al., 1991). It is not known if this is a general characteristic shared with other members of the genus.
Calliandra is considered to be native to the New World and it is well represented from the southwestern states of the USA, through Central America and into the warmer parts of South America, including Argentina and Chile. Some species are also found in India, Madagascar and West Africa
(Dalziel, 1937; Uphof, 1968; Allen and Allen, 1981; Mabberley, 1987). C. calothyrsus is widely commercialized in Indonesia and it was introduced into the region from Guatemala in 1936 (NRC, 1983; Baggio and Heuveldop, 1984).
Fodder characteristics
The species which has attracted most attention for its capacity to produce both fuelwood and foliage for either green manure or fodder is C. calothyrsus, a small tree which grows to about 10 m in height. It is of Central and South American origin, occurring naturally in moist, tropical regions up to an altitude of some 1500 m. While it grows up to 2000 m in Kenya, production is limited at this altitude, probably by the low temperatures (Lowry and Macklin, 1988). It appears to do best with annual rainfall in the range of 2000-4000 mm (NRC, 1983). While it will grow in areas that receive 700-1000 mm rain/year, productivity is reduced by low rainfall (Akkasaeng et al., 1989). It is evergreen in humid environments but will shed its leaves during a long, dry season. Under conditions of severe drought, young stems and branches may die back, but they usually regrow when the rains return. Mature branches become brittle and may be easily broken by animals, although this is not a problem where judicious cutting is practiced.
The palatability of the foliage appears to be variable but it is accepted, at least in limited quantities and when mixed with other feeds, by most livestock, including sheep, goats, cattle and water buffalo (Brewbaker et al., 1983; NRC, 1983; Baggio and Heuveldop, 1984). It has been classified as unpalatable to rabbits, although they consumed significant amounts of it when mixed with grass and herbaceous leaves. Foliage of other fodder trees such as Leucaena leucocephala and Albizia falcataria were eaten in much larger quantities (Raharjo and Cheeke, 1985). In Java, Calliandra leaf meal is used at levels of up to 5% in diets for chickens (Panjaitan, 1988).
Until recently, most plantings of C. calothyrsus were based on seed from a limited number of provenances from poorly documented sites in Guatemala, Costa Rica and Honduras. In 1990, a programme was initiated by the Oxford Forestry Institute (based in UK) to collect seed over the entire natural range of this, and several other closely related Calliandra spp. The objective was to provide material both for testing in multi-locational trials for future breeding activities. Seed distribution for wide international assessment of provenances was expected to commence in 1993 (Macqueen, 1991; Pottinger, 1992), but was actually initiated in 1992 and greatly expanded in the following year (D. Macqueen, personal communication). Assessment is in the early stages but results of this work will become available in due course.
In Rwanda, infertile soils of pH 4.3 and 4.8, with low levels of aluminium saturation and rainfall of 1166-1564 mm/year, allowed satisfactory growth of C. calothyrsus in the absence of fertilizers, although there was a small positive interaction between applications of lime at 750 kg/ha and farmyard manure at rates of between 2.5 and 10.0 t/ha (Yamoah et al., 1989). On acid (pH about 5), infertile soils with either high or low levels of aluminium saturation in both Australia and Indonesia, this species showed considerable promise when harvested at intervals of about three months, outyielding both Leucaena leucocephala and Gliricidia septum, particularly in the absence of fertilizer application (Bray et al., 1989; Palmer et al., 1989).

Table 1 - Proximate and fibre
analyses of Calliandra calothyrsus
These results indicate the ability of C. calothyrsus to tolerate highly acidic and infertile soils.
In a pot experiment with soil limed at varying rates to adjust the acidity from pH 4.3 (no lime) to a maximum of pH 8.0, best growth of C. calothyrsus was obtained in the range of pH 6-8 (Hu et al., 1983). Netera et al. (1992) showed considerable differences in growth characteristics between two lines of C. calothyrsus in a pot experiment on an infertile oxisol (pH 5 and aluminium saturation about 60%) from West Java. While these observations may indicate considerable genetic variation within the genus, the authors suggested that the line which showed poor adaptation to acid soil may be a species other than C. calothyrsus. They pointed out the need to detail the source of seed fully when quoting experimental results. This, however, is very rarely done at present.
In Java and Sumatra, at four sites between sea level and 920 m altitude, with soil pH at 5.6 6.9 and rainfall at 1200-3600 mm/year, C. calothyrsus produced higher yields of leaf material than seven other leguminous trees at the highest site, and was the second-best producer at two other sites (Panjaitan et al., 1989). The in vitro dry matter digestibility as measured by the cellulase digestibility technique was relatively low however, ranging from 24.8% in a dry environment to 51.1% at a site with good rains in all months of the year. Despite the poor levels of digestibility, it was rated as the most agronomically adaptable of the species tested, comparing well with Leucaena leucocephala, Gliricidia septum, Sesbania spp. and Albizia falcataria in terms of growth and resistance to pests and diseases.
On the island of Sumba (Indonesia), at altitudes of 500-1000 m and with shallow, clay loam soils, a series of leguminous trees was tested as potential alternatives to Leucaena leucocephala where psyllid (Heteropsylla cubana) attacks were a recurring problem. With annual rainfall of about 1000 mm spread mainly over a 5 month wet season, C. calothyrsus was considered to be useful, even though growth slowed dramatically with the onset of flowering, about 2 months into the dry season. With slightly higher rainfall (1200-1500 mm), C. tetragona (syn. Zapoteca tetragona) was found to be a more productive species but no information is available regarding animal production (Rourke and Suardika, 1990). In contrast to this report, NRC (1983) noted that C. tetragona was generally slower growing and less satisfactory than C. calothyrsus in Indonesia. More work is required to clarify the situation regarding the relative merits, and perhaps even the delineation of these and other species of Calliandra.
There are few results available for C. calothyrsus planted specifically for animal production, but in an alley cropping experiment in western Samoa, cutting at 1.5 m every 4-5 months gave slightly higher yields of leaf dry matter over 13 months (12.9 t/ha) than cutting at 1 m (10.4 t/ha). Total biomass production (21.4 t/ha) was the same from both treatments and cutting height had no effect on the nutrient content of the foliage (Tekle-Haimanot et al., 1991) In a separate 4-year alley cropping experiment in the same region, on a moderately fertile soil receiving annual rainfall of about 3000 mm, yields of taro (Colocasia esculenta) were slightly better with Gliricidia septum hedges than when C. calothyrsus formed the hedges, even though annual applications of mulch from the Calliandra (9.6 t/ha DM) averaged 11% more than with the Gliricidia (Rosecrance et al., 1992). There were no significant differences between species in terms of their effects on the physical or chemical properties of soil. On a relatively infertile soil of pH 6.3 and annual rainfall of 1250 mm in southwestern Nigerta, C. calothyrsus produced some 6 t DM/ha in non-wood prunings (leaves and small twigs) in four clippings per year. This provided some 200 kg/ha of nitrogen for use by plants as the mulch decomposed (Gichuru and Kang, 1989). The growth and performance of the Calliandra was comparable to that of Leucaena leucocephala as a hedge for alley cropping. It therefore represents a real alternative for regions where insect problems threaten the use of the better known species.
Crude protein (CP) contents of C. calothyrsus are often quoted at 20-22% (e.g. Ahn et al., 1989) and some typical analyses are quoted in Table 1. In Sumatra, however, on an acid ultisol, Blair et al. (1988) obtained much lower CP values of 13.7%. The differences could be due to soil fertility, but they may be due to differences in accessions, or even species, of Calliandra since there is a degree of taxonomic confusion within the genus (Netera et al., 1992). In common with several other tree genera on the acid soils of South Sumatra, the foliage of C. calothyrsus contained adequate levels of potassium, calcium and magnesium for animal production (Blair et al., 1988), although it was poor in both phosphorus (0.11%) and sodium (0.01%). At three sites in Java, Jakarta and North Sumatra, however, phosphorus levels of 0.16-0.19% were recorded by Panjaitan (1987), levels which appear to be marginal for beef cattle (McDowell et al., 1983).
When wilted, fresh leaves and edible stems of C. calothyrsus were suspended for 48 hours in intraruminal nylon bags in steers maintained on a diet of Elephant grass (Pennisetum purpureum) and concentrates, DM disappearance was 51.0% (Mahyuddin et al., 1988). When the material was dried, DM disappearance was reduced to 31.5-37.2% depending on the drying technique (either at differing temperatures in an oven, in the sun, or by freeze drying). Similarly, reductions in digestibility as a result of drying were measured using both in vitro pepsin-cellulase and rumen fluid-pepsin techniques. Oven-drying the fodder resulted in decreases in both total phenolic compounds and condensed tannins compared with freeze drying (Akin et al., 1989) and this was reflected in an increase in in sacco nitrogen digestibility in goats. Unfortunately, this work did not evaluate fresh, unwilled fodder.
In a recent, short-duration experiment using Merino wethers of about 27 kg liveweight, C. calothyrsus was fed as the sole dietary component. Daily DM consumption of fresh material (35% DM) was 5.9 kg/100 kg metabolic body weight and material wilted by a forced draught at ambient temperature (95% DM) was consumed at 3.7 kg/100 kg of metabolic body weight. These levels of voluntary intake are high, even for small ruminants, and indicate high acceptability of the foliage. In Droughtmaster steers, in sacco digestibility of wilted and oven-dried material was similar, and inferior to that of fresh foliage. Digestibility of fresh material was 60%, reducing to about 30% after only 6 hours of forced draught wilting (Palmer and Schlink, 1992).
These studies would appear to explain some of the conflicting reports regarding the acceptability, digestibility and feeding value of C. calothyrsus. The results quoted above suggest that in order to obtain maximum benefit from the feeding of this species, it should be used as a protein supplement for poorer quality roughage, either browsed direct, or offered in limited quantities to animals as soon as possible after cutting, at which time it will compare well with other tree foliage in terms of feeding quality. The species appears to have little potential for use as dried leaf meal, at least where ruminant animals are concerned, because of the rapid loss of digestibility after cutting.
Anti-nutritive factors
Toxic compounds such as alkaloids and cyanogenic glycosides do not occur in Calliandra spp. but a range of insecticidal, non-protein, sulphur-containing amino acids and rare imino acids have been found in the seeds and seedlings of a number of species. Only the amino acids have been isolated from mature leaves (Bleecker and Romeo, 1981, 1983; Romeo et al., 1983). These compounds are known to have toxic effects against a range of leaf and seed-eating insects, and it is suggested that the seeds and tender young leaves, those parts which need most protection from the point of view of survival of the species, benefit from a dual defence mechanism comprised of both amino and imino acids. Mature leaves, which have had time to develop physical attributes such as toughness and waxiness, require only the protection conferred by the imino acids (Romeo and Swain, 1986). There are no reports of adverse effects of these compounds on higher animals.
In common with many other trees, Calliandra spp. appear to be characterized by high levels of phenolic compounds, including tannins. NRC (1983) reported that the level of vanillin-reacting compounds (condensed tannins) in leaves of C. calothyrsus seemed to be 1-3%, but in a more recent evaluation, the leaves of a single sample of this species contained 18.18% total phenolics. These compounds included condensed tannins of 11.07% when measured by the Vanillin-HCI technique, or 2.05% by the Butanol-HCI method (Akin et al., 1989). Such high tannin levels were reflected by in sacco nitrogen digestibility of only 35.9% in goats, a figure which confirmed in vitro dry matter digestibility of leaves of the same species of 35.43% (Baggio and Heuveldop, 1984). As noted above, the digestibility of C. calothyrsus decreases rapidly as the foliage wilts. The cause of this is not known, but it may be associated with the presence of unusually high levels of tannins (Palmer and Schlink, 1992).
Management
Calliandra spp. generally produce good-quality seed, but despite the presence of protective, insecticidal chemicals, the seed crop may be susceptible to destruction by insect pests. In Kenya, the rose flower beetle (Pachnoda ephippiata) and related species (P. ancticollis and P. viridana) feed on fruits, flowers and foliage of a number of trees and have been blamed for poor seed harvests of C. calothyrsus. Infestation appears to be aggravated by long spells of dry weather (Kaudia, 1990). In the Philippines, the larvae of an unidentified tussock moth have been reported attacking the flowers of C. calothyrsus. The moth has so far caused only minor damage but has the potential to become a significant pest (Braze, 1991). In both cases the tree was a recent introduction. It may be that chemical defences that evolved in the Americas proved to be ineffective against indigenous insect pests.
Seed of Calliandra spp. stores well, particularly under refrigeration, but this is not usually necessary as many species have long flowering periods and produce seed for most of the year. While the seed appears to germinate well without scarification, it has been suggested that germination may be speeded up by treatment with hot water, or by soaking in cold water. Recommended practices include pouring boiling water over the seed and allowing it to cool and soak for 24 hours (NRC, 1983), or immersion for 5 minutes in hot water, followed by soaking in water at room temperature. The latter method produced a germination rate of 77.3% in Costa Rica (Baggio and Heuveldop, 1984). Small seedlots may be scarified by nicking the coat of each individual seed.
As well as establishment either by direct seeding or by transplanting seedlings raised in nurseries Calliandra spp. reproduce readily by vegetative means. Both large stakes and young coppice with at least two nodes can be successfully rooted.
Establishment is assisted by the use of indole butyric acid rooting hormone (Tomaneng, 1991), but readily achieved without chemical assistance.
After an initial establishment period, Calliandra spp. tolerate frequent, regular cutting. Established C. calothyrsus trees were pruned at intervals of 3 months over a 4-year alley cropping trial on an acid, infertile soil in West Sumatra. The trees remained productive under this management and overall survival rate was 97%, compared with 61% for Paraserianthes falcataria Gliricidia septum had a survival rate of about 34% and only persisted where high levels of lime (2-4 t, had been applied to reduce the levels of aluminium saturation (Dierolf and Yost, 1989).
On an alluvial soil of pH 6 in South Sulawesi (Indonesia), trees were established from seedlings and allowed to grow in a pure stand for a year before being cut back to a height of 1 m. Subsequent regular cutting at 3-month intervals over an 18-month period resulted in 92% survival of trees of C. calothyrsus, while cutting every 6 weeks led to 81% survival. Leaf yield, particularly during the wet season, increased with increasing tree density in the range of 500040 000 trees/ha and also with the longer intervals between harvests. Overall, the cumulative yield of leaves was 8.5 t DM/ha/year (Ella et al., 1989). The yield of Guinea grass (Panicum maximum cv. Riversdale), which was planted under the trees at the conclusion of the work described above was reduced by the longer harvesting interval, particularly at higher tree densities. Overall, the understorey of grass produced yields of 7.2 t DM/ha/ year and the combination of grass and trees outyielded either component when grown in a pure stand. The presence of the grass did not change the behaviour of the trees. It was suggested that the grass and the trees obtained their moisture from different levels in the soil profile, and that during the dry season, the trees provided the grass with some protection from drying winds (Ella and Blair, 1989; Ella et al., 1991 b).
Initial work on highly acid, volcanic soil in Costa Rica, in an area with annual rainfall of 2600 mm, suggested that trees of C. calothyrsus planted at spacings of 25-50 cm could be used as hedges to subdivide grazing areas. Rapid tree growth and high survival rates in the field produced plants of over 120 cm in height within 4 months of planting from seedlings raised in the nursery (Baggio and Heuveldop, 1984). In general terms, the yield of leaf from fodder trees at subsequent harvests is positively related to the age of the tree at the first cutting, at least within the first year of two of its life. The yield differences are much less in C. calothyrsus than other tree species such as Leucaena leucocephala and Gliricidia septum, however, and are almost negligible between 13 and 17 months (Ella et al., 1991a). This would suggest that the first cutting of either hedges or individual trees could be carried out when convenient after a limited establishment period, without greatly influencing subsequent fodder production.
A range of fungal diseases including rusts, dieback, leaf blotch and pink disease have been recorded on Calliandra spp. in a number of countries.
They seldom become serious problems however, except where rough coppicing may allow infection to kill weakened tree stumps, particularly if they are cut too close to ground level. Scales and trunk borers are sometimes found on branches and stems, and loopers may damage some leaves. In plant nurseries, snails and rats occasionally destroy tightly packed seedlings, but in general terms Calliandra spp. appear to show good resistance to pests and diseases and control measures are rarely necessary (Lowry and Macklin, 1988; Lenne, 1992).
Changes occur in the digestibility of the foliage in the first few hours after cutting which, together with the loss of leaves in response to severe moisture stress, highlights lunitations in the selection of suitable animal management systems involving Calliandra spp. (Palmer and Schlink, 1992). Direct browsing or rotational harvesting programmes based on daily cutting and rapid feeding of small quantities of fodder, are favoured because of the ability of the plant to withstand frequent defoliation. Alternative sources of good quality fodder must be provided for the dry season however, when tree growth slows and there is a danger of losing the feed as a result of leaf shedding.
High levels of tannins in the foliage probably cause the relatively slow rate of microbial decomposition where Calliandra leaves are applied to the soil. This characteristic suggests that the species may be more effective as a mulch than as green manure in alley cropping systems (Salazar and Palm 1987).
Alternative uses
Members of the genus Calliandra are used for a variety of purposes in a number of countries (Uphof, 1968; Allen and Allen, 1981; NRC, 1983).
The wood is hard, heavy (specific gravity 0.5-0.8), strong but tending to become brittle at maturity, medium-textured, and easy to work, although it is not highly lustrous. Several species, including C. formosa (syn. Zapoteca formosa), are used for small carpentry, implement handles and frames in tropical America.
The species are valued for fuel since they often exhibit high growth rates and respond well to coppicing.
The wood of C. calothyrsus dries rapidly and burns well, giving off 4600 kcal/kg of heat. It also makes good charcoal: in Indonesia it can produce annual yields of up to 14 t/ha of charcoal with a calorific value of 7200 kcal/kg. Some areas have been coppiced annually for more than 20 years, producing 35-65 m3/ha/year of dry wood (NAS, 1979; Lowry and Macklin, 1988). The wood pulp is easily bleached and is used as a filler at levels of up to 10% in the making of paper.
C. calothyrsus is often used in reafforestation in Indonesia, to combat soil erosion and to recover and improve the fertility of bare and degraded land. It is attracting increasing attention for use as the hedge component in alley cropping systems (NRC, 1983; Gichuru and Kang, 1989). Although use is made of many naturally occurring Calliandra spp., only C. calothyrsus and C. tetragona (syn. Zapoteca tetragona) appear to have been planted for forestry purposes.
The bark of C. anomala (syn. C. grandiflora) is used in tanning in Central America, and the root is used to retard fermentation in the making of alcoholic beverages. In the past, the bark of C. houstoniana (syn. C. houstonii) has been marketed in Europe as a substitute for quinine, and as an antiperiodic.
Extracts from the roots of C. grandiflora have been used in Mexico to treat eye diseases, diarrhoea and indigestion, while in West Africa the dried and powdered leaves of C. portoricensis were sniffed to relieve headaches (Dalziel, 1937).
The flowering period of many species is long, and the nectar is attractive to honey bees. The honey from C. calothyrsus has a pleasing, bittersweet flavour and annual honey yields of 1 t/ha of tree plantation have been estimated in Java. The tree has also been shown experimentally to be a suitable host for the lac insect Kerria lacca, a valuable producer of shellac.
Most members of the genus produce attractive foliage and colourful flowers. Species such as C. grandifora, C. inaequilatera, C. surinamensis and C. tweedii are particularly valued for their large flowers and these, and many other species are planted as individual trees or hedges in gardens and along roadsides and plot boundaries in many tropical countries.
Immature seeds of C. calothyrsus are eaten raw or fried by indigenous people in parts of Mexico (Macqueen, 1993).
References and further reading
AHN, J.H., ROBERTSON, B.M., ELLIOTT, R. GUTTERIDGE, R.C. and FORD, C.W. (1989) Quality assessment of tropical browse legumes: tannin content and protein degradation. Animal Feed Science and Technology, 27:147-156.
AKKASAENG, R., GUTTERIDGE, R.C. and WANAPAT, M. (1989) Evaluation of trees and shrubs for forage and fuelwood in northeast Thailand. International Tree Crops Journal, 5:209-220.
ALLEN, O.N. and ALLEN, E.K. (1981) The Leguminosue: A Source Book of Characteristics, Uses and Nodulation. Madison, USA: University of Wisconsin Press.
BAGGIO, A. and HEWELDOP, J. (1984) Initial performance of Calliandra calothyrsus Meissm. in live fences for the production of biomass. Agroforestry Systems, 2:19-29.
BLAIR, G.J., PANJAITAN, M., IVORY, D.A., PALMER, B. and SUDJADI, M. (1988) An evaluation of tree legumes on an acid ultisol in South Sumat Indonesia. Journal of Agricultural Science, 111:435-441.
BLEECKER, A.B. and ROMEO, J.T. (1981) 2,4-trans 4,5-trans-4,5-dihydroxypipecolic acid and cic-5 hydroxypipecolic acid from leaves of Calliandra angustifolia and sap of C. confusa. Phytochemistry, 20:1845-1846.
BLEECKER, A.B. and ROMEO, J.T. (1983) 2,4-cis-4, 5cis-4, 5-dihydroxypipecolic acid: a naturally occurring imino acid from Calliandra pittieri. Phytochemistry, 22:1025-1026.
BRAY, R.A., PALMER, B. and IBRAHIM, T. (1989A) multisite trial of shrub legumes on acid soils. Nitrogen Fixing Tree Research Reports, 7:7-8.
BRAZA, R.D. (1991) Insects damaging Cailliandra calothyrsus in the Philippines. Nitrogen Fixing Tree Research Reports, 9:38-39.
BREWBAKER, J.L., HALLIDAY, J. and LYMAN, J. (1983) Economically important nitrogen fixing tree species. Nitrogen Fixing Tree Research Reports, 1:35-40.
BREWBAKER, J.L., WILLERS, K.B. and MACKLIN, B. (1990) Nitrogen fixing trees: validation and prioritization. Nitrogen Fixing Tree Research Reports, 8:8-16.
DALZIEL, J.M. (1937) The Useful Plants of West Tropical Africa. London: Crown Agents for the Colonies.
DIEROLF, T.S. and YOST, R.S. (1989) Survival rates of three tree species in a four-year-old alley cropping trial. Nitrogen Fixing Tree Research Reports, 7:12-13.
ELLA, A. and BLAIR, G.J. (1989) Effect of tree density and cutting frequency on the production of four tree legumes and understorey grass. Nitrogen Fixing Tree Research Reports, 7:14-16.
ELLA, A., BLAIR, G.J. and STUR, W.W. (1991a) Effect of age of forage tree legumes at first cutting on subsequent production. Tropical Grasslands, 25:275280.
ELLA, A., JACOBSEN, C., STUR, W.W. and BLAIR, G. (1989) Effect of planting density and cutting frequency on the productivity of four tree legumes. Tropical Grasslands, 23:28-34.
ELLA, A., STUR, W.W., BLAIR G.J. and JACOBSEN, C.N. (199lb) Effect of plant density and cutting frequency on the yield of four tree legumes and interplanted Panicum maximum cv. Riversdale. Tropical Grasslands, 25:281-286.
EVANS, D.O. and ROTAR, I.P. (1987) Productivity of Sesbania species. Tropical Agriculture (Trinidad), 64:193-200.
GICHURU, M.P. and KANG, B.T. (1989) Calliandra calothyrsus (Meissn.) in an alley cropping system with sequentially cropped maize and cowpea in southwestern Nigeria. Agroforestry Systems, 9:191-203.
GUTTERIDGE, R.C. (1990) Agronomic evaluation of tree and shrub species in southeast Queensland. Tropical Grasslands, 24:29-34.
HU, T.W., CHENG, W.E. and SHEN, T.A. (1983) Growth of the seedlings of four leguminous tree species in relation to soil pH in a pot test. Nitrogen Fixing Tree Research Reports, 1:24-25.
KAUDIA, A. (1990) Report of an insect pest on Calliandra calothyrsus (Meissn.) in Kenya. Nitrogen Fixing Tree Research Reports, 8:126.
LOWRY, J.B. and MACKLIN, W. (1988) Calliandra calothyrsus: an Indonesian favourite goes pantropical. NFT Highlights, 88-02. Waimanalo, Hawaii: Nitrogen Fixing Tree Association.
LENNE, J.M. (1992) Diseases of multipurpose woody legumes in the tropics: a review. Nitrogen Fixing Tree Research Reports, 10:13-29.
MABBERLEY, D.J. (1987) The Plant Book. New York: Cambridge University Press.
MACQUEEN, D.J. (1991) Exploration and collection of Calliandra calothyrsus as a foundation for future genetic improvement. Nitrogen Fixing Tree Research Reports, 9:96-98.
MACQUEEN, D.J. (1993) Calliandra Series Racemosae Taxonomic Information; OFI Seed Collections; Trial Design. Oxford: Oxford Forestry Institute.
McDOWELL, L.R., CONRAD, J.H., ELLIS, G.L. and LOOSLI, J.K. (1983) Minerals for Grazing Ruminants in Tropical Regions. Gainesville, USA: University of Florida/US Agency for International Development.
MAHYUDDIN, P., LITTLE, D.A. and LOWRY, J.B. (1988) Drying treatment drastically affects feed evaluation and feed quality with certain tropical forage species. Animal Feed Science and Technology, 22:69-78.
NAS (1979) Tropical Legumes: Resources for the Future. Washington, D.C., USA: National Academy Press.
NETERA, S.N., PALMER, B. and BRAY, R.A. (1992) Differential response to phosphorus and lime of two accessions of Calliandra calothyrsus on an acid soil. Nitrogen Fixing Tree Research Reports, 10:62-65.
NRC (1983) Calliandra: A Versatile Small Tree for the Humid Tropics. Washington, D.C., USA: National Academy Press.
PALMER, B., BRAY, R.A., TATANG IBRAHIM and FIJLLOON, M.G. (1989) Shrub legumes for acid soils. In: Management of Acid Soils in the Humid Tropics of Asia. CRASSWELL, E.T. and PUSHPARAJAH, E. (eds). Monograph No.13. Canberra, Australia: Australian Centre of International Agricultural Research (ACIAR).
PALMER, B. and SCHLINK, A.C. (1992) The effect of drying on the intake and rate of digestion of the shrub legume Calliandra calothyrsus. Tropical Grasslands, 26:89-93.
PANJAITAN, M. (1988) Nutritive value of tree legumes introduced in Indonesia. IARD Journal, 10:73-80.
PANJAITAN, M., JESSOP, R. and STUR, W.W. (1989) Evaluation of tree legume species at different agroclimatic zones of Indonesia. pp. 945-946. In: Proceedings, XVI International Grassland Congress, Nice, France, October 1989. Versailles, France: Association Fran�ais pour la Production Fourrag�re.
POTTINGER, A.J. (1992) Nitrogen fixing tree research networks co-ordinated by the Oxford Forestry Institute. Nitrogen Fixing Tree Research Reports, 10:7-12.
RAHARJO, Y.C. and CHEEKE, P.R. (1985) Palatability of tropical tree legume forage to rabbits. Nitrogen Fixing Tree Research Reports, 3:31-32.
ROMEO, J.T. and SWAIN, L.A. (1986) Persistence of non-protein seed amino acid S-(�-carboxyethyl) cysteine in young leaves of Calliandra rubescens: ecological implications. Journal of Chemical Ecology, 12:2089-2096.
ROMEO, J.T., SWAIN, L.A. and BLEECKER, A.B. (1983) cis-4-hydroxypipecolic acid and 2,1-cis-4,5trans-4, 5-dihydroxypipecolic acid from Calliandra. Phytochemistry, 22:1615-1617.
ROSECRANCE, R.C., ROGERS, S. and TOFINGA, M. (1992) Effects of alley cropped Calliandra calothyrsus and Gliricidia septum on weed growth, soil properties, and taro yields in western Samoa. Agroforestry Systems, 19:57-66.
ROURKE, M.K. and SUARDIKA, I.P.P. (1990) Onfarm species trials to investigate several NFT species for use in terraced hedgerows. Nitrogen Fixing Tree Research Reports, 8:37-39.
SALAZAR, A. and PALM, C.A. (1987) Screening of leguminous trees for alley cropping on acid soils of the humid tropics. pp. 61-67. In: Gliricidia septum (Jacq.) Walp.: Management and Improvement. Proceedings, Workshop, CATIE, Turrialba, Costa Rica, June 1987. Special Pub. 87-01. Hawaii, USA: Nitrogen Fixing Tree Association.
SHELTON, H.M., LOWRY, J. B., GUTTERIDGE, R.C., BRAY, R.A. and WILDIN J.H. (1991) Sustaining productive pastures in the tropics: 7. Tree and shrub legumes in improved pastures. Tropical Grasslands, 25:119-128.
TEKLE-HAIMANOT, A., WEERARATNA, C.W. and DOKU, E.V. (1991) Evaluation of three tree legume species in western Samoa. Nitrogen Fixing Tree Research Reports, 9:71-74.
TOMANENG, A.A. (1991) Production of Calliandra seedlings from young coppice cuttings. Nitrogen Fixing Tree Research Reports, 9:99.
UPHOF, J.C. Th. (1968) Dictionary of Economic Plants. 2nd edn. New York: Verlay von J. Cramer.
YAMOAH, C., GROSZ, R. and NIZEYIMANA, E. (1989) Early growth of alley shrubs in the Highland region of Rwanda. Agroforestry Systems, 9:171-184.
Other titles in the Use of Trees by Livestock series include:
1 Prosopis
2 Acacia
3 Gliricidia
4 Anti-nutritive
factors
5 Quercus
6 Cassia
7
Ficus
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Use of Trees by Livestock (NRI, 1994, 160 p.)
Use of Trees by Livestock: Erythrina
R.T. Paterson
Acknowledgements
© Crown copyright 1994
The Natural Resources Institute (NRI) is an internationally recognized centre of expertise on the natural resources sector in developing countries. It forms an integral part of the British Government's overseas aid programme. Its principal aim is to alleviate poverty and hardship in developing countries by increasing the productivity of their renewable natural resources. NRI's main fields of expertise are resource assessment and farming systems, integrated pest management, food science and crop utilization.
NRI carries out research and surveys; develops pilot-scale plant, machinery and processes; identifies, prepares, manages and executes projects; provides advice and training; and publishes scientific and development material.
Short extracts of material from this publication may be reproduced in any non-advertising, nonprofit-making context provided that the source is acknowledged as follows: Paterson, R.T. (1994) Use of Trees by Livestock 9: Erythrina. Chatham, UK: Natural Resources Institute.
Permission for commercial reproduction should be sought from:
The Head, Publishing and Publicity Services, Natural Resources Institute, Central Avenue, Chatham Maritime, Kent, ME4 4TB, United Kingdom.
No charge is made for single copies of this publication sent to governmental and educational establishments, research institutions and non-profitmaking organizations working in countries eligible for British Government Aid. Free copies cannot normally be addressed to individuals by name but only under their official titles. Please quote LG9 when ordering.
Natural Resources Institute ISBN: 0 85954 381-1 Price £2.00
This booklet is printed on paper produced using the residues from sugar-cane processing.
Foreword
The importance of trees and shrubs in the feeding of animals in the tropics and sub-tropics has long been recognized by livestock owners. In arid areas where the growth of herbaceous plants is limited by lack of moisture, leaves and edible twigs of trees and shrubs can constitute well over 50% of the biomass production of rangeland. At high altitudes, tree foliage may provide over 50% of the feed available to ruminants in the dry season, branches being harvested and carried to the animals. Even in regions of higher rainfall where grass supplies the major proportion of the dry matter eaten by ruminants, tree leaves and fruits can form an important constituent of the diet, particularly for small ruminants.
In the last two decades interest in the planting of trees as a source of feed for livestock has been encouraged by workers in research and development, but in contrast to the hundreds of indigenous species which are used as fodder, attention has focussed on a limited number of introduced species. Thus there are many publications reporting the chemical composition of Leucaena leucocephala leaves and suggesting management strategies for utilization of the tree for fodder, but it is more difficult to find information on alternative genera which might be equally, or more, appropriate.
The aim of this series of publications is to bring together published information on selected genera of trees which have the potential to increase the supply of fodder for ruminants. Each booklet summarizes published information on the fodder characteristics and nutritive value of one genus, with recommendations on management strategies, where available. Further, since the leaves of woody species frequently contain secondary compounds which may have an antinutritional, or toxic, effect, a separate booklet summarizes the effects of a number of these compounds. It is hoped that the booklets will provide useful resource material for students, research and extension workers, interested in promoting the use of trees as a source of fodder for ruminants.
Further copies of this booklet or others in the series can be obtained by writing to Publishing and Publicity Services at the Natural Resources Institute.
Margaret Gill
Livestock Production
Programme
Genus Erythrina
Family LEGUMINOSAE
Subfamily PAPILIONOIDEAE
Tribe
PHASEOLEAE
Subtribe ERYTHRINAE
Principal species
Erythrina abyssinica
Erythrina berteroana
Erythrina
costaricensis
Erythrina crista-galli
Erythrina edulis
Erythrina
fusca (syn. E. glauca)
Erythrina variegata (syn. E. indica)
Erythrina
poeppigiana (syn. E. micropteryx)
Erythrina tahitensis (syn. E sandwicensis,
E. monosperma, E. montana)
Erythrina senegalensis
Erythrina subumbrans
(syn. E. Iithosperma)
Main common names
Coral trees (India, southern Africa)
Madre arbor
Central
and
Madre de cacao
Por�South
America
Summary
Erythrina is a genus of some 108 species of shrubs and trees which are distributed widely throughout the tropics and sub-tropics. Several species thrive on waterlogged, or poorly drained, acid soils which are inhospitable to most legumes. While they have long been used in Africa, Asia and the Americas as ornamentals, living fence posts, supports for vine crops, green manure and as shade trees in plantations of coffee, their use as fodder trees has attracted much less attention until recent years. With increased awareness of the role of trees in animal production interest has focussed on the genus, despite concern over the effects on livestock of a range of alkaloids and flavonoids which are known to occur in the foliage. There are no reports of toxicity in domestic livestock. Indeed, use is made of many Erythrina spp. to treat a range of ailments in both human and veterinary medicine.
Erythrina is rich in crude protein but the digestibility of the foliage is only about 50%. Many species are well accepted by livestock, particularly small ruminants, and there is considerable potential for using them as dietary supplements. They may be of particular use in areas where there is a lack of other high-quality fodder species due to soil acidity and lack of drainage.
Description and distribution
Erythrina is a genus of some 108 species of shrubs and small to medium-sized trees (to about 20 m in height), which are often armed with blunt, conical thorns or sharp, recurved prickles which may occur on the trunks, young branches, petioles, leaf midribs and main veins. The leaves vary in shape, but are often deltoid or rhomboid. They are pinnately trifoliate and the terminal leaflet is often the largest. The lateral leaflets may be asymmetric. Erythrina spp. tend to shed their leaves under the influence of moisture stress, and where this happens the flowers often appear before, or with, the first new leaves at the start of the growing season. The flowers are usually red but may also be pink, orange or yellow. They are large and attractive, giving rise to the use of many species as ornamental plants. The seeds are commonly red to orange, or brown in colour, with a contrasting black (or sometimes white) patch at the point of attachment. They are carried, 2-14 at a time, in long, flattened or cylindrical, dehiscent pods with deep constrictions between the segments (Allen and Allen, 1981; Coates Palgrave, 1983).
Erythrina spp. occur over a wide range of natural habitats including open forest, dry brush and scrub, river banks, swamps and coastal regions. One species, E. tahitensis, is reported to occur at altitudes of up to 2900 m (Allen and Allen, 1981), and E. edulis is commonly found between 1800 and 2500 m. Maximum altitudes for E. fusca and E. poeppigiana are considered to be 1400 and 1700 m respectively, while E. tahitensis occurs naturally only at altitudes up to about 600 m. Those species which are adapted to higher altitudes show some frost tolerance, and most are susceptible to damage by fire (Coates Palgrave, 1983). In the Americas, species such as E. fusca are often found on the most acid and infertile of soils, and they thrive in waterlogged and poorly drained areas where fast-growing, nitrogen-fixing trees seldom prosper (Preston and Murgeitio, 1987). In Ethiopia, E. burana occurs in a range of situations, including shallow, swampy bogs and dry, rocky hills with slightly alkaline (pH 7.1-7.3), sandy or gravelly soils (Teketay, 1990).
Distribution of the genus is pantropical, with some 70 species found in the Americas, 32 in Africa, 18 in Asia and 3 in Australia and Argentina (Allen and Allen, 1981). See Krukoff and Barneby (1974) for a conspectus of the species.
Fodder characteristics
Brewbaker (1989) suggested that Erythrina spp. had little forage potential in the African context. There are a number of reports in the literature (e.g. Jama et al., 1989; Tarawali, 1991) which indicate that introductions of a range of species have not always met with success because of agronomic and biotic problems, such as damping off of seedlings and damage caused by stem borers. Nevertheless, E. abyssinica is used in the Wolayata region of southern Ethiopia for dry season fodder, live fences and shade for coffee (Lazier and Mengistu, 1984) and its potential as a fodder tree is thought to be worthy of further study (Larbi et al., 1993). E. variegata (syn. E. indica) is one of a total of 13 fedder shrubs and trees found to be widely used in ruminant feeding systems in Asia (Chen et al., 1992), while E. arborescens is planted by Nepalese farmers in areas of 1000-1500 m altitude (Joshi, 1992). E. fusca and E. poeppigiana are of growing importance to livestock in Central and South America (Beer, 1980; Preston and Murgeitio, 1987; Preston, 1992; Kass et al., 1992).
In the humid tropics of Costa Rica, E. poeppigiana interplanted with King grass (Pennisetum purpureum x P. typhoides hybrid) in blocks and pruned three or four times per year, gave dry matter yields of tree fadder ranging from 6.4 t/ha/year at a density of 1667 trees/ha to 11.3 t/ha/year with 3333 trees/ha (Benavides et al., 1989). When planted at high density in fertilized, pure stands, E. berteroana produced 19.4 t/ha/year of DM in three harvests (CATIE, 1989). In a similar climatic region, where several Erythrina spp., including the native species E. berteroana, E. fusca and E. globocalyx, together with E. poeppigiana of South American origin, are commonly used as living fences (Sauer, 1979), fouryear-old trees of E. berteroana, established from large stakes, yielded 319 kg DM of leaves and stems from 100 m of fence line (169 trees) after eight months growth (Budowski et al., 1985). With typical planting arrangements where trees are grown at spacings ranging from 1-3 m, annual DM yields in the range of 1.8-3.0 t/km of fenceline were obtained (CATIE, 1989).
Table 1 - Proximate and fibreanalyses
of Erythrina spp.
Many Erythrina spp. are well accepted by many classes of livestock. The leaves of E. variegata are eaten by cattle in India (CSIR, 1986), while both the leaves and the young flowers of E. sigmoidea are relished by zebu cattle in Cameroon (Audru, 1980). Goats consumed leaves of E. poeppigiana at a daily level of up to 3.2 kg DM/100 kg liveweight (Samur, 1984), while sheep and goats consumed E. abyssinica at levels of 3.0 and 2.8 kg OM (organic matter)/100 kg of metabolic weight (Larbi et al., 1993). The fruit of E. edulis is consumed by pigs in Colombia (Preston and Murgeitio, 1987), while in Indonesia (Raharjo and Cheeke, 1985), rabbits ate up to 23% of their diet as chopped leaves of E. subumbrans (syn. E. Iithosperma).
The nutritive value of Erythrina spp. is generally held to be good, although published data are scarce. Representative values are presented in Table 1. Crude protein (CP) contents of 30.�, 11.3 and 13.8% respectively have been reported for the leaves, stem and bark of E. poeppigiana (Preston and Murgeitio, 1987). The CP content of dried and ground leaf meal of this species, averaged over seven individual evaluations in Costa Rica, was 26.1% (calculated from Kass et al., 1992). The CP content of leaves and stems of E. berteroana (at 8 months growth) were similar at 26.3 and 8.1% respectively (Budowski et al., 1985).
The in vitro dry matter digestibility of the leaves and stems of E. poeppigiana appears to be somewhat variable, but moderate and in the range of 44-53% (Samur, 1984; Preston and Murgeitio, 1987; Rodriguez et al., 1987; Benavides et al., 1989; Kass et al., 1992). The variation could be due to differing analytical techniques, or to varying amounts of stem in the samples, although Salazar and Vasquez (1988) observed considerable genetic variation in morphology between different provenances of this species in Costa Rica. This variability may also be reflected in the nutritional value of the foliage. The bark had a remarkably high level of digestibility which Preston and Murgeitio (1987) reported to be 78%.
The published literature is sparse but available data suggest that some African and Asian species of Erythrina may be lower in protein content and higher in digestibility, at least in small ruminants, than the better known American species. Chopped and wilted leaves of E. abyssinica from Ethiopia had CP levels of 20.6% with in vivo DM digestibility of about 65% in both sheep and goats. This species was considered to show promise as a cheap source of protein (Larbi et al., 1993). The protein content of a single sample of E. arborescens from Nepal was only 16.9% (Josh), 1992), but this figure could be artificially low if an unusually high proportion of edible stem was included in the sample.
Dried leaves of E. variegata showed in sacco DM disappearance of only 36% after incubation in cows of 12-72 hours, but 60% of the CP was degraded (Huq and Saadullah, 1987). These results suggest that Erythrina leaves may be more effective when used to supplement poor quality pastures than would be predicted on the basis of laboratory evaluation of dry matter digestibility alone.
In Costa Rica, where it is common to supplement dairy cattle with concentrates based on soyabeans, the replacement of the concentrate with E. poeppigiana foliage resulted in a linear decrease in daily weight gains in young heifers and in the milk yields of cows. Dry matter intake of grass was also reduced. The growth rate of grazing steers on unsupplemented diets, however, increased when the steers were fed low levels of E. cocleata fodder, and growth rates were further improved by the addition of an energy source such as green bananas or molasses. Work with rice bran suggested the need to supply bypass nutrients (both energy and protein) to obtain high milk yields when Erythrina fodder was offered to dairy cattle as the main source of dietary nitrogen. It was concluded that under lowland humid conditions, the direct benefits to milk production from the use of Erythrina forage would be small where alternative, conventional concentrates were available. There may, however, be important indirect benefits such as increased carrying capacity of pasture as a result of fodder substitution, and improved nutrient recycling within the grazing area. The financial benefits would be site-specific, depending on relative costs and returns. In subhumid regions, supplementation of poor quality roughages with tree foliage may be an attractive method of improving animal productivity and profitability (Kass et al., 1992).
The foliage of Erythrina spp. appears to be particularly attractive to small ruminants. Milk yields of dairy goats fed on a basal diet of King grass (Pennisetum purpureum) and reject bananas were increased linearly by 84-156% when the animals were given leaves of E. poeppigiana at daily levels of 0.5-1.5 kg DM/100 kg liveweight. The percentage increase was similar for goats with both high and low initial milk yields (Esnaola and Rios, 1990). Animals consumed 16% more foliage of E. poeppigiann than Gliricidin sepium when offered under similar conditions, and daily milk yields increased by some 15% (Rodriguez et al., 1987). The tree fodder produced lower increases in yields than commercial concentrates with the same level of CP, suggesting that the tree protein was of lower nutritional value than conventional supplements. Using costs and returns from Costa Rica, economic analyses strongly favoured the use of Erythrina fodder over concentrates based on soyabeans (Kass et al., 1992).
The weight gains of young, castrated male sheep and goats fed over an 80-day period on a basal diet of Napier grass (Penissetum purpureum) almost doubled when the diet was supplemented with chopped, wilted leaves of E. abyssinica at rates of up to 1 kg/head/day (Larbi et al., 1993). Again, the response to increasing levels of the tree fodder appeared to be linear.
In general, with small ruminants fed on King or Napier grass, there was a partial substitution of Erythrina for the basal diet, but total dry matter intake increased as a result of the feeding of the tree fodder (Kass et al., 1992).
The high nutritional value of the genus in terms of digestible crude protein is sufficient to justify its use as a protein supplement to improve the feeding value of diets based on poor-quality grass or roughage.
Anti-nutritive factors
A recent bibliography of Erythrina (Agishi and Tothill, 1991) contained a total of 344 entries and no fewer than 97 papers referred to alkaloids and medicinal uses, although few of these were concerned with anti-nutritive factors. Alkaloids which have been isolated from members of the genus include hypaphroine, erythroidine (Allen and Allen, 1981), erysotine, erythratidine and epi-erythratidine (Chawla et al., 1988). The presence of physiologically active alkaloids is considered to be characteristic of the genus, although flavonoids including the isoflavones auriculasin and scandenone have also been found in some species (Nkengfack et al., 1989). Some of these flavonoids have been shown to have anti-fungal (Maillard et al., 1978) and anti-microbial (Biyiti et al., 1988) properties, but their effects on higher animals are unknown.
All Erythrina spp. so far tested have yielded alkaloids which have effects on small laboratory animals (e.g. frogs) that are similar to poisoning by curare, the poison used by some Indians in South America on their arrows. The symptoms include drowsiness and muscular paralysis of the neck, extremities and diaphragm. When death occurs, it usually results from respiratory failure. The most potent species are E. buchii, E. coralloides, E. cristagalli, E. eggersii, E. macrophylla, E. mexicana, E. Ianata subsp. occidentalis (syn. E. occidentalis) and E. suberosa (Allen and Allen, 1981). Toxic effects have been produced by the injection of leaf extracts containing concentrated alkaloids into the bloodstream of the test animals, but Coates Palgrave (1983) thought it unlikely that ingestion of potentially poisonous seeds of E. abyssinica and E. caffra would have any adverse effects on humans.
The leaves of E. subumbrans are palatable to rabbits, although they are less well accepted than those of some other trees such as Leucaena leucocephala and Albizia falcataria (Raharjo and Cheeke, 1985). It has been reported, however, that some Erythri, Zn spp. can cause sterility and even death in rabbits (Martin, 1984). Reports of toxicity in larger domestic animals have not been found.
Erythrina spp. are generally lower in soluble polyphenolic compounds and lignin, and thus decompose more rapidly in the soil, than other tropical legumes such as l?Inga edulis and Cajanus cajan (Salazar and Palm, 1987). It has been suggested (Palm and Sanchez, 1990) that polyphenolics bind to nitrogenous compounds in the leaves to form substances that are resistant to decomposition. This may be analogous to the way in which high levels of tannins in the foliage of some browse species protect them from digestion when eaten by livestock. The low level of polyphenolics in Erythrina is a positive characteristic from the point of view of animal nutrition.
Management
The seeds of most Erythrina spp. are fertile and show little sign of dormancy (Rao and Singh, 1987), some, such as E. abyssinica may be hard seeded, a condition which can be rectified by scarification in either hot water or concentrated sulphuric acid (Laurent and Chamshama, 1987). Mechanical scarification is recommended for E. tahitensis (Powell and Nakao, 1992). Vegetative propagation is often favoured since most species root easily (Napier, 1988). Stakes of a minimum 2.5 cm diameter and 30 cm long will usually gro, but planting is often carried out using larger stakes of 10 cm diameter and some 2-2.5 m in length. Root formation, which usually takes place readily, can be further stimulated by the use of hormonal preparations, or by stripping the bark from the lower end of the part to be buried in the soil (Teketay, 1990). This end is cut obliquely and the upper end is protected against both rain and desiccation by the application of tar or lime. In a number of countries, apicormic shoots (large shoots that grow vertically upwards from horizontal branches) of E. costaricensis, E. variegata, E. poeppigiana and E. senegalensis have proved to be particularly successful as planting material (Jolin and Torquebiau, 1992).
Planting densities in the range of 1200-10 000 trees/ha are common. Lower densities are used where groups of trees are intended to provide shade and browse within a pasture, while at the upper end of the range, the trees would form a pure stand with very little understorey. When large stakes are used, growth is rapid and the first harvest can often take place within some 6-8 months of planting (Preston and Murgeitio, 1987).
Erythrina spp. form large, spherical nodules, which tend to be clustered on the central tap root. They are infected promiscuously by Bradyrhizobium spp. of the cowpea group, and therefore rarely require inoculation. The list of species which have been shown to nodulate includes all of those which are commonly used in animal production (Allen and Allen, 1981). At the low tree densities employed as shade for coffee and cacao, annual nitrogen fixation rates are 12-40 kg/ha (Budowski et al., 1986). Active mycorrhizal associations are formed which are valuable in soils with low phosphorus contents (Powell and Nakao, 1992).
Grass-based pasture growing under seven-yearold trees of E. poeppigiana established as shade for grazing animals (about 60 trees/ha) was compared with similar pasture growing either without shade or under the tree species Cordia alliodora, Albizia saman (syn. Samanea saman) or Gliricidia septum in Costa Rica, grass yields were similar in all cases, but the fibre content of the grass was higher in the unshaded areas. The CP content of the grass was highest under E. poeppigiana, and lowest under C. alliodora (Daccarett and Blydenstein, 1968). While direct measurements of the nitrogen fixing capacity of Erythrina spp. are lacking for pasture systems, these data would suggest that it is at least comparable to that of G. septum, a much-used fodder tree in tropical regions.
Frequent cutting appears to be advantageous when Erythrina spp. are grown for animal production. In a coffee plantation in the humid tropics of Costa Rica (2600 mm annual rainfall) where E. poeppigiana was planted for shade at a density of 280 trees/ha, the legume was cut at intervals of 4, 6 or 12 months. The most frequent pollarding produced the lowest total yield of dry matter (woody stems, edible stems and leaves), but the highest yield of edible stems and leaves. With frequent cutting, the edible fraction was some 64% of the total biomass production, and contained about 85% of the total crude protein. The total amount of nitrogen in the cut material was 170 kg/ha at three cuts per year, compared with 230 kg/ha under more frequent cutting systems (Rodriguez, 1985; Russo and Budowski, 1986).
In a study conducted over two years in a similar area of Costa Rica E. poeppigiana was planted in association with King grass and neither weeded nor fertilized. The trees were established as large stakes at either 1667 or 3333 trees/ha, and cut three or four times a year. When harvested at varying intervals, on attainment of a height of 2 m, grass production was not reduced by the presence of the trees at either density, but the crude protein content (average over two years) was increased from 4.7 to 6.1%. The higher tree density almost doubled the yield of tree fodder. More frequent cutting of the trees slightly increased tree leaf yield, but had little effect on crude protein content, which averaged 26.3% for the leaf and 11.7% for the stem (Benavides et al., 1989). Since the stems of the trees are of limited feeding value, and leaves may be shed in response to dry weather, livestock production would benefit from management under frequent cutting, although the growth rate of the trees, as influenced by climatic, edaphic and biotic factors, would govern the optimum interval between harvests. A heavy pruning of Erythrina spp. not only promotes the yield of leaves and prevents loss of feed resources resulting from leaf fall, it also appears to prevent the early senescence that is sometimes observed in trees after a few years of active growth (Seibert, 1987).
Alternative uses
The alternative uses of the genus Erythrina are described by a number of authors including: Dalziel (1937); Uphof (1968); Wickens (1980); Allen and Allen (1981); Coates Palgrave (1983) Budowski et al., (1986); and CSIR (1986).
Young leaves and tender shoots of species such as E. variegata, E. fuseda and E. rubrinenvia are eaten raw or cooked in India, Indonesia and Central America. Flowers and buds of E. rubrinervia are also consumed as a vegetable. The seeds of E. edulis and E. variegata are boiled and roasted for human consumption in parts of Colombia and India, although those of E. variegata may be poisonous when raw.
Decoctions of the leaves, bark and roots of many species, including E. corallodendron, E. herbacea and E. senegalensis, are used for medicinal purposes in the treatment of wounds and ailments such as jaundice, dysentery, bronchitis and venereal diseases, and to alleviate toothache. The powdered bark of E. senegalensis is administered in water to horses as a diuretic, while E. variegata appears to have anthelmintic properties.
Because of their alkaloid content and their curare like properties, the seeds, stems and bark of many species of Erythrina find uses as poisons for fish, while the powdered seeds of E. herbacea are used to control rats in Mexico. Water extracts of the leaves of E. variegata are highly toxic to the nematodes Meloidogyne incognita and Tylenchorhynchus mashhoodi, offering potential benefits to small farmers for cheap, organic plant pest control (Mohanty and Das, 1988).
The brightly coloured seeds of several species, including E. abyssinica and E. caffra, are strung as beads for decoration, and a fibre derived from the bark of E. senegalensis is made into scented necklaces and bracelets.
The wood is soft, light (specific gravity about 0.25), spongy and usually pale in colour. While rather woolly to work by machine, it may be carved for toys, statuettes and assorted small items including marimbas (type of musical instrument), drums, ladles, jars, stools and packing cases. The wood of E. tahitensis is particularly light and has been used in Hawaii as floats for fish nets and surf boards. It is a favourite canoe wood throughout Polynesia. In general, the wood has little value as either fuelwood or charcoal. The twigs of E. senegalensis may be used as feather dusters.
The bark of E. suberosa is used on the Indian subcontinent in the manufacture of cork plugs and insulation boards. The wood, bark and ash of this species are also used in dyeing.
Several species such as E. berteroana, E. cochleata, E. costaricensis, E. fusca, E. poeppigiana and E. subumbrans (syn. E. Iithosperma), make excellent live fences, shade and green manure trees in plantations of tea, coffee and cocoa, and support trees for vine crops such as peppers. E. americana is amongst the most common live fences in the Tabasco region of Mexico (Vera Castillo, 1987). Many species, including E. abyssinica, E. caffra, E. crista-galli and E. variegata are valued as ornamental shrubs and trees by virtue of their attractive, brightly coloured flowers.
References and further reading
AGISHI, E.C. and TOTHILL, J.C. (1991) A Bibliography on the Brozuse Trees Calliandra, Chamaecytisus and Erythrina. Plant Science Division Working Document No. 5. Addis Ababa, Ethiopia: International Livestock Centre for Africa (ILCA).
ALLEN, O.N. and ALLEN, E.K. (1981) The Leguminosae: A Source Book of Characteristics, Uses and Nodulation. Madison, USA: The University of Wisconsin Press.
AUDRU, J. (1980) Ligneous and subligneous forage and fruit trees in the Guinean zone: prospects for utilization in animal production. In: LE HOUEROU, H.N. (ed.). Browse in Africa The Current State of Knowledge. Addis Ababa, Ethiopia: International Livestock Centre for Africa (ILCA).
BEER, J. (1980) Erythrina poeppigiana con pasto: un estudio de cave. In: SOMARRIBA, E. (ed.). Investigaci�n Agroforestal del Proyecto UNU/CATIE 1979-1987. Informe T�cnico No.131. Turrialba, Costa Rica: Centro Agron�mico de Investigaci�n y Ense�anza (CATIE).
BENAVIDES, J.E., RODRIGUEZ, R.A. and BOREL, R. (1989) Producci�n y calidad del forraje de pasto king grass (Pennisetum purpureum x P. typhoides) y por� (Erythrina poeppigiana) sembrados en associaci�n. In: XANDE, A. and ALEXANDRE, G. (eds). Paturages et Alimentation en Zone Tropicale Humide. Paris: Institut National de la Recherche Agronomique (INRA).
BIYITI, L., PESANDO, D. and PUISEUX-DAO, S. (1988) Antimicrobial activity of two flavonones isolated from the Cameroonian plant Erythrina sigmoidea. Planta Medica, 54:126-128.
BREWBAKER, J.I. (1989) Nitrogen fixing trees for fodder and browse in Africa. In: KANG, B.T. and REYNOLDS, L. (eds). Alley Farming in the Humid and Subhumid Tropics. Workshop held at Ibadan, Nigeria, March 1986. Ottawa, Canada: IDRC.
BUDOWSKI, G., RUSSO, R. and GLOVER, N. (1986) Erythrinas provide beauty and more. NFT Highlights, 86-02. Hawaii, USA: Nitrogen Fixing Tree Association.
BUDOWSKI, G., RUSSO, R.O. and MORA, E. (1985) Productividad de una cerca viva de Erythrina berteroana Urban en Turrialba, Costa Rica. Turrialba, 35(1):83-86.
CATIE (1989) Informe Anual 1988/89. Turrialba, Costa Rica: Centro Agron�mico de Investigaci�n y Ense�anza (CATIE).
CHAWLA, A.S., KRISHNAN, T.R., JACKSON, A.H. and SCALABRIN, D.A. (1988) Alkaloidal constituents of Erythrina variegata bark. Planta Medica, 54:526-528.
CHEN, C.P., HALIM, R.A. and CHIN, F.Y. (1992) Fodder trees and shrubs in range and farming systems of the Asian and Pacific region. In: SPEEDY, A. and PUGLIESE, P. (eds). Legume Trees and Other Fodder Trees as Protein Sources for Livestock. FAO Animal Production and Health Paper 102. Rome: Food and Agriculture Organization (FAO).
COATES PALGRAVE, K. (1983) Trees of Southern Africa. 2nd edn. Capetown: C. Struik.
CSIR (1986) The Useful Plants of India. New Delhi: Publications and Information Directorate, Council of Scientific and Industrial Research (CSIR).
DACCARETT, M. and BLYDENSTEIN, J. (1968) The influence of leguminous and non-leguminous trees on the forage that grows beneath them. Turrialba, 18(4):405-408.
DALZIEL, J.M. (1937) The Useful Plants of West Troyical Africa. London: Crown Agents for the Colonies.
ESNAOLA, M.A. and RIOS, C. (1990) Hojas de Por� (Erythrina poeppigiana) como suplemento proteico pare cabras lactantes. Livestock Research for Rural Dez~elopment, 2(1):24-33.
HUQ, M.A. and SAADULLAH, M. (1987) Ruminal dry matter and nitrogen degradability of common tree leaves and forages in Bangladesh. Indian Journal of Animal Nutrition, 4(1):44-47.
JAMA, B., NAIR, P.K.R. and KURIRA, P.W. (1989) Comparative growth performance of some multipurpose trees and shrubs grown at Machakos, Kenya. Agroforestry Systems, 9:17-27.
JOLIN, D. and TORQUEBIAU, E. (1992) Large cuttings: a jump start for tree planting. Agroforestry Today, 4(4):15-16.
JOSHI, N.P. (1992) Use of fodder trees and shrubs as protein supplements to ruminants and as a means of soil stabilization: the Nepalese case. In: SPEEDY, A. and PUGLIESE, P. (eds). Legume Trees and Other Fodder Trees as Protein Sources for Livestock. FAO Animal Production and Health Paper 102. Rome: Food and Agriculture Organization (FAO).
KASS, M., BENAVIDES, J., ROMERO, F. and PEZO, D. (1992) Lessons from main feeding experiments conducted at CATIE using fodder trees as part of the N-ration. In: SPEEDY, A. and PUGLIESE, P. (eds). Legume Trees and Other Fodder Trees as Protein Sources for Livestock. FAO Animal Production and Health Paper 102. Rome: Food and Agriculture Organization (FAO).
KRUKOFF, B. A. and BARNEBY, R.C. (1974) Conspectus of species of the genus Erythrina. Lloydia, 37:332-459.
LARBI, A., THOMAS, D. and HANSON, J. (1993) Forage potential of Erythrina abyssinica: intake, digestibility and growth rates for stall-fed sheep and goats in southern Ethiopia. Agroforestry Systems, 21.
LAURENT, N. and CHAMSHAMA, S.A.O. (1987) Studies on the germination of Erythrina abyssinica and Juniperus procera. International Tree Crops Journal, 4:291-298.
LAZIER, J. and MENGISTU, S. (1984) Erythrina: a genus with browse potential. Plant Genetic Resources Germplasm Newsletter, 5:20-22. Add is Ababa, Ethiopia: International Livestock Centre for Africa (ILCA).
MAILLARD, M., GUPTA, M.P. and HOSTETTMAN, K. (1978) A new antifungal prenylated flavonone from Erythrina berteroana. Journal of Medicinal Plant Research, 53:563-564.
MARTIN, F.W. (1984) Edible leaves from nitrogen fixing trees. Nitrogen Fixing Tree Research Reports, 2:57-58.
McDOWELL, L.R., CONRAD, J.H., THOMAS, J.E. and HARRIS, L.E. (1974) Latin American Tables of Feed Composition. Gainesville, USA: University of Florida.
MOHANTY, K.C. and DAS, S.N. (1988) Nematicidal properties of Erythrina indica against Meloidogyne incognita and Tylenchorhynchus mashhoodi. Indian Journal of Nematology, 18:138.
NAPIER, I.A. (1988) Vegetative propagation of fodder trees. Preliminary research results and practical recommendations. Banko Janakari, 2(1):21-30.
NKENGFACK, A.E., SANSON, D.R., FOMUM, Z.T. and TEMPESTA, M.S. (1989) 8-Prenylluteone, a prenylated isoflavone from Erythrina eriotriocha. Phytochemistry, 28:2522-2526.
PALM, C.A. and SANCHEZ, P.A. (1990) Decomposition and nutrient release patterns of the leaves of three tropical legumes. Biotropica, 22:330338.
POWELL, M.H. and NAKAO, P.L. (1992) Erythrina sandwicensis: unique Hawaiian NFT. NFT Highlights, 92-06. Hawaii, USA: Nitrogen Fixing Tree Association.
PRESTON, T.R. (1992) The role of multi-purpose trees in integrated farming systems for the wet tropics. In: SPEEDY, A. and PUGLIESE, P. (eds). Legume Trees and Other Fodder Trees as Protein Sources for Livestock. FAO Animal Production and Health Paper 102. Rome: Food and Agriculture Organization (FAO).
PRESTON, T.R. and MURGEITIO, E. (1987) Tree and shrub legumes as protein sources for livestock. In: WALMSLEY, D. (ed.). Forage Legumes and Other Local Protein Sources as Substitutes for Imported Protein Meals. Proceedings, CTA/CARDI seminar held at Kingston, Jamaica, December 1987. St. Augustine, Trinidad and Tobago: Caribbean Agricultural Research and Development Institute (CARDI).
RAHARJO, Y.C. and CHEEKE, P.R. (1985) Palatability of tropical tree legume forage to rabbits. Nitrogen Fixing Tree Research Reports, 3:31-32.
RAO, P.B. and SINGH, S.P. (1987) Effect of light quality on seed germination of certain woody species of Kumaun Himalaya. Acta Botanica Indica, 15:247-254.
RODRIGUEZ, F. (1985) Biomass production of intersown Erythrina poeppigiana (Warpers) O.F. Cook and King grass (Pennisetum purpureum x typhoides), as a function of the sowing density and pruning frequency of E. poeppigiana. Actividades en Turrialba, 13(3):20.
RODRIGUEZ, Z., BENAVIDES, J.E., CHAVES, C. and SANCHEZ, G. (1987) Producci�n de leche de cabras estabuladas alimentadas con follaje de madero negro (Gliricidia septum) y de por� (Erythrina poeppigiana) y suplementadas con platano pelipita (Musa sp. cv. Pelipita). pp. 212-216. In: Gliricidia septum (Jacg.) Walp.: Management and Improvement. Proceedings, Workshop held at CATIE, Turrialba, Costa Rica, June 1987. Special Pub. 87-01. Hawaii, USA: Nitrogen Fixing Tree Association.
RUSSO, R.O. and BUDOWSKI, G. (1986) Effect of pollarding frequency on biomass of Erythrina poeppigiana as a coffee shade tree. Agroforestry Systems, 4:145-162.
SALAZAR, A. and PALM, C.A. (1987) Screening of leguminous trees for alley cropping on acid soils of the humid tropics. pp. 61-67. In: Gliricidia septum (Jacq.) Walp.: Management and Improvement. Proceedings, Workshop held at CATIE, Turrialba, Costa Rica, June, 1987. Special Pub. 87-01. Hawaii, USA: Nitrogen Fixing Tree Association.
SALAZAR, R. and VASQUEZ, M.S. (1988) Variaci�n gen�tica en ocho procedencias de Erythrina poeppigiana en Costa Rica. Turrialba, 38(2):71-81.
SAMUR, C. (1984) Producci�n de Leche de Cabras Alimentadas con King Grass (Pennisetum purpureum) y Por� (Erythrina poeppigiana) Suplementadas con Fruto de Banano (Muse sp. cv. Cavendish). MSc. thesis, Turrialba, Costa Rica: Universidad de Costa Rica/Centro Agron�mico Tropical de Investigaci�n y Ense�anza (CATIE).
SAWER, J.D. (1979) Living fences in Costa Rican agriculture. Turrialba, 29(4):255-261.
SEIBERT, B. (1987) Pepper on living poles of Gliricidia septum and Erythrina lithospermum in East Kalimantan. pp. 111-118. In: Gliricidia septum (Jacg.) Walp.: Management and Improvement. Proceedings, Workshop held at CATIE, Turrialba, Costa Rica, June 1987. Special Pub. 87-01. Hawaii, USA: Nitrogen Fixing Tree Association.
TARAWALI, S.A. (1991) The potential of selected nitrogen fixing trees in Central Nigeria: growth parameters. Nitrogen Fixing Tree Research Reports, 9:27-30.
TEKETAY, D. (1990) Erythrina burana: promising multipurpose tree from Ethiopia. Agroforestry Today, 2(4):13.
UPHOF, J.C.T. (1968) Dictionary of Economic Plants. 2nd edn. New York: Verlay von J. Cramer.
VERA CASTILLO, G. (1987) Estado actual de la investigaci�n en Gliricidia septum (Jacq.) Walp. en Mexico. pp. 14-19. In: Gliricidia septum (Jacq.) Walp.: Management and Improvement. Proceedings, Workshop held at CATIE, Turrialba, Costa Rica, June, 1987. Special Pub. 87-01. Hawaii, USA: Nitrogen Fixing Tree Association.
WICKENS, G.E. (1980) Alternative uses of browse species. In: LE HOUEROU, H.N. (ed.). Browse in Africa: The Current State of Knowledge. Addis Ababa, Ethiopia: International Livestock Centre for Africa (ILCA).
Other titles in the Use of Trees by Livestock series include:
1. Prosopis
2. Acacia
3. Gliricidia
4. Anti-nutritive
factors
5. Quercus
6. Cassia
7. Ficus
8.
Calliandra